Основные генетические параметры белкового полиморфизма в природных популяциях сиговых рыб.
Для исследованных выборок сиговых рыб были рассчитаны основные параметры генетической изменчивости — величины уровня полиморфизма и средней гетерозиготности (табл. 6.8). Необходимо отметить, что уровень полиморфизма находится в положительной корреляции С величиной исследованной выборки, поэтому в случае небольших выборок значения полиморфизма могут оказаться искусственно заниженными, поэтому индексы относительной гетерозиготности более адекватно отражают величину генетической изменчивости, наблюдающуюся в популяции.
Таблица 6.8
Уровни генетической изменчивости в природных популяциях сиговых рыб

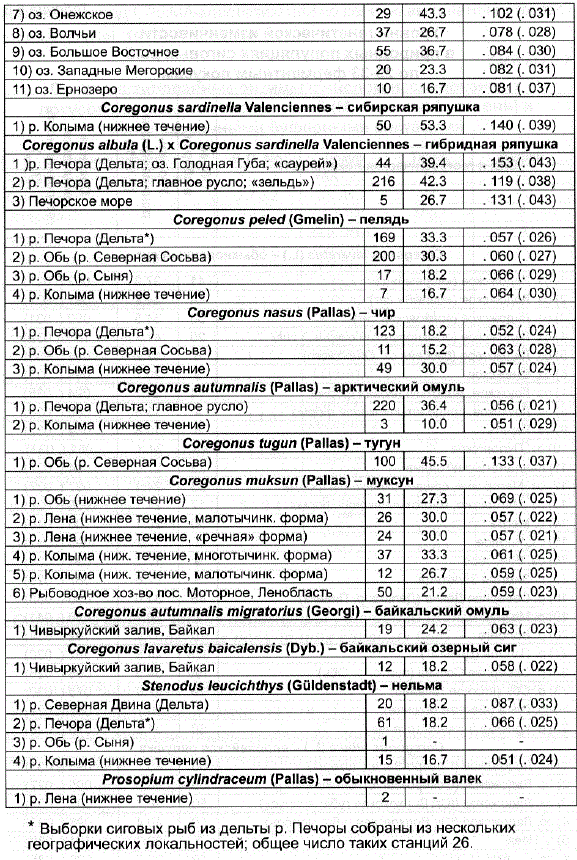
* Выборки сиговых рыб из дельты р. Печоры собраны из нескольких географических локальностей; общее число таких станций 26.
Наши данные показали, что виды сиговых рыб в значительной степени различаются по этим параметрам, что подтверждает сделанный ранее вывод о существенной гетерогенности сем. Coregonidae по величинам наследственной изменчивости (Кирпичников, 1987; Ермоленко, 1989а). К числу рыб с наибольшими значениями средней гетерозиготности и полиморфизма относятся ряпушки, что совпадает с результатами, полученными ранее (Vuorinen, 1984; Перелыгин, 1988; Ермоленко, 1989а). Максимальные значения обоих параметров были отмечены у сибирской ряпушки из низовьев Колымы (H 53,3 и P=0,140), а также у двух форм Печорской ряпушки (H 39,4-42,3 и P=0,119-0,153), что объясняется их гибридным происхождением: полиморфизм двух исходных предковых таксонов – Coregonus albula и Coregonus sardinella – суммировался у их дериватов - у двух локальных форм —» зельди» и «саурея». Отметим, что и колымская ряпушка по ряду генетических маркеров демонстрирует спектры полос, характерные для гибридов, что косвенным образом может указывать на зону гибридизации разных приледниковых рас в пределах вида Coregonus sardinella.
Близкие к максимальным значения Рин, получены для тугуна (P=45,5 ин 0,133), который так же, как и типичная ряпушка, является самым мелким видом р. Coregonus. Возможно, что высокая генетическая изменчиВость имеет для тугуна адаптивное значение в связи с большой удельной Поверхностью тела и эвритермностью этих рыб (Пирожников и др., 1975). - К высоко полиморфным видам относится сиг. Значительная генетическая вариабельность вида с. lavаrеtus объясняется биологией этого вида. Как известно, обыкновенный сиг, имеющий, как и ряпушки, циркумполярное распространение, в большей степени, чем другие виды сем. Coregonidae, способен к освоению разнообразных экологических ниш; в результате широкой адаптивной радиации Сига в пределах данного вида появились многочисленные экологические и морфологические формы (Берг, 1948; Решетников, 1980). По-видимому, потенциальная возможность быстрой адаптации в гетерогенных условиях окружающей среды требует поддержания достаточно высоких параметров ферментного полиморфизма у этих видов. Генетическая лабильность позволяет виду в условиях разных биотопов с успехом приспосабливаться к действию многих факторов окружающей среды, которые лимитируют распространение других видов сиговых.
Значительным биологическим полиморфизмом обыкновенного сига объясняется пуществование полярных взглядов по вопропу ето внутривидовой систематики: часть исследователей признает наличие одного полиморфного вида с несколькими (от 6 до 31) подвидами (Берг, 1948; Правдин, 1954; Шапошникова, 1976, 1977; Решетников 1980,1995). По мнению других, С. lavаrеtus представлен несколькими видами (Thienemann, 1928; Jarvi, 1928,1943; Wagler, 1937; Svardson, 1952,1957,1979; Dottrens, 1959; Himberg, 1970; Китаев, 1983), причем количество этих видов только в водоемах Европы может достигать 44 единиц (Коttеlаt, Freyhof, 2007).
Ярким примером противоположной биологической стратегии может служить вид с. nasus. Чир, являющийся типичным бентофагом, имеет весьма суженный спектр питания, популяциям этого вида свойственно достаточно ограниченное пространственное распространение с приуроченностью к низовьям рек и предустьевым участкам. По-видимому, именно «экологической забуференностью» чира объясняются невысокие значения генетического разнообразия, выявленные нами в популяциях вида. Схожие выводы на основе ранее полученных результатов были сделаны в работах А. Б. Локшиной (1980) и Л.Н. Ермоленко (1989а).
Низким уровнем полиморфизма и невысокой гетерозиготностью характеризуются и другие специализированные виды сиГовых рыб. Так, у представителя хищных сиговых рыб – нельмы — процент полиморфных локусов оказался на уровне P=16,7-18,2, хотя относительная гетерозиготность колебалась в более широком диапазоне - Ho0,051-0,087. Уровень относительной гетерозиготности печорской популяции арктического oмyля оказался равным н 0,056, похожая величина индекса выявлена у колымского омуля – на 0,051. Как известно, арктический омуль ведет полупроходной образ жизни, и, возможно, невысокие значения генетической изменчивости обусловлены адаптацией этого вида к строгому жизненному циклу, связанной со сменой солености среды обитания. У представителей байкальской сиговой фауны, различающихся между собой по . кормовой специализации, — байкальского омуля (планктофаг) и байкальского озерного сига (эврифаг) — индексы полиморфизма и гетерозиготности также оказались невысокими и не превысили величины P=24,2 и на 0,063. ,
Интересно отметить факт невысоких значений индексов генетического разнообразия (прежде всего относительной гетерозиготности) муксуна при сравнении его с обыкновенным сигом, учитывая филогенетическую близость двух этих видов. В самом деле, если у вида с. lavаrеtus относительная гетерозиготность только пяти из 25 исследованных популяций была ниже величины H 0,070, то все шесть проанализированных популяций вида С. muksun характеризовались индексами, не превышающими это значение. Популяции пеляди рек Печоры, Оби и Колымы в целом характеризуются невысокими значениями индексов генетической изменчивости и между собой по индексу относительной гетерозиготности различаются незначительно: Не 0,057 – 0,066.
Таблица 6.9 Распределение генетического разнообразия по иерархическим уровням в популяциях ряпушки, пеляди и муксуна*

*Исследованные популяции сиговых рыб относятся к водосборам следующих морей: ряпушка – Балтийское, Белое, Баренцево, Восточно-Сибирское – всего 15 популяций; сиг – Балтийское, Белое, Баренцево, Карское, Лаптевых, ВосточноСибирское — всего 20 популяций; пелядь – Баренцево, Карское, ВосточноСибирское – всего 5 популяций; муксун — Карское, Лаптевых, Восточно-Сибирское - всего 5 популяций.**В случае с муксуном анализу подвергались симпатрически обитающие разнотычинковые формы рыб.
Для четырех видов сиговых рыб был проведен анализ распределения выявленного генетического разнообразия по иерархическим уровням. Так, для 15 популяций комплекса ряпушек на географическом пространстве от Балтийского моря до Восточно-Сибирского доля межпопуляционного разнообразия не превысила 17%, причем почти 10% общей изменчивости приходилось на межрегиональную составляющую (табл. 6.9). На внутрипопуляционную изменчивость 20 популяций сига С. lavаrеtus, представляющих водоемы от Балтики до р. Колымы, приходилось 88,3% общей генетической вариабельности, при этом межпопуляционное разнообразие подразделялось на различия между крупными водосборами (6,0%) и дифференциацию локальных популяций в пределах этих водосборов (5,7%).
Таким образом, оценки генетической изменчивости на внутрипопуляционном уровне у двух рассматриваемых видов сиговых рыб с циркумполярным ареалом сопоставимы между собой и подтверждают справедливость сделанного ранее заключения о менее сложной внутривидовой организации рыб сем. Coregonidae по сравнению, например, с представителями сем. Salmonidae (Андрияшева, 1996, 2009; Титов, 2007). Существенные различия между сигами и ряпушками по доле межрегиональной Компоненты изменчивости, по-видимому, объясняются тем, что процесс интрогрессивной гибридизации между дериватами различных приледниКовых рас в постледниковое время мог иметь значительную пространственную глубину. Таким образом, индекс различия между регионами, представленными исследованными популяциями сигов, неизбежно будет снижаться. С другой стороны, в перечень проанализированных популяций ряпушек были включены как представители чистых европейской и сибирской ряпушек, так и выборки печорской ряпушки, гибридной по своему происхождению, что, напротив, степень межрегиональных различий для комплекса ряпушек увеличивает.
В отличие от закономерностей, выявленных при распределении генетического разнообразия в популяциях сига и ряпушки, те же оценки для популяций пеляди и муксуна оказались смещенными в сторону увеличения доли внутрипопуляционной компоненты, составляя 92,5 и 94,0% соответственно. Более низкие величины межпопуляционной составляющей, минимальные при сравнении на уровнях ниже регионального, по-видимому, могут объясняться как слабой популяционной подразделенностью всех видов сиговых рыб, так и (в отличие от ряпушки и сига) . сравнительно недавним распространением пеляди и муксуна по северу Евразии из единого центра расселения, находящегося на территории южной или центральной Сибири. Небезынтересно отметить, что на долю генетических различий между разнотычинковыми формами муксуна из рек Лены и Колымы приходится всего 2,1% от общей генетической вариабельности популяций этого вида.
Сравнительное исследование генетического полиморфизма популяций позволило оценить родственные связи между таксонами сиговых рыб, относящихся к группам разного уровня организации. Так, проведенные межвидовые генетические сопоставления рыб сем. Coregonidae показали, что выборки сиговых агрегируются в несколько дискретных генетических групп, составленных отдельными видами семейства. Картина взаимоотношений между этими группами представлена в виде дендрограммы, построенной на основании индексов генетических расстояний (рис. 6.2).

Рис. 6.2. UPGMA дендрограмма генетических различий между исследованнымивидами сиговых рыб
Из «хороших» видов, традиционно выделяемых систематиками, найболее близкими друг к другу оказались сиг и муксун (D-0,010), а также байкальский омуль и байкальский озерный сиг (D-0,011). Подобный невысокий уровень различий обычно характерен для географически разобщенных популяций одного вида, но не для разных видов: например, уровень генетических расстояний между исследованными нами популяциями вида С. lavаrеtus на территории Северо-Запада России и Западной Сибири в целом невысок, но в ряде случаев может превышать величину D-0,050 (Sendek, 2004). Поэтому, будучи включенными в совместный анализ с сигами, популяции муксуна не образуют на дендрограмме отдельного кластера, как правило, «растворяясь» среди популяций сига, представляющих разные районы своего ареала. Результаты ранее проведенных сравнительных исследований сигa и муксуна позволили авторам работ (Politov et al., 2002; Балдина и др., 2008) сделать заключение о несоответствии морфологических и генетических критериев при сопоставлении сигa и муксуна и, возможно, об определенной искусственности выделения муксуна в отдельный вид. Но Тем не менее оценки генетической дифференциации двух названных видов, основанные преимущественно на суммарных величинах индексов сходства или различия, далеко не в полной мере могут отражать эволюционные взаимоотношения между этими таксонами. В подобных случаях тесных родственных отношений значительно более информативным может оказаться анализ распространения в популяциях видов отдельных качественных признаков, находящихся под строгим генетичеСким контролем. Так, например, во многих популяциях вида С. lavаrеtus с достаточно высокой частотой (p>0,1) обнаруживается аллель IDDH*f, который не встречается в симпатрически обитающих вместе с сигом популяциях муксуна на географическом пространстве от p. Оби до р. Колымы. Отсутствие данного маркерного признака в генофонде муксуна предполагает наличие достаточно мощного репродуктивного барьера между сигом и муксуном, величина которого сопоставима с силой изолирующих механизмов, ограждающих целостность генофондов других сиговых рыб от взаимного «размывания» при спонтанной гибридизации. Процент появления природных гибридов между сигом и муксуном в целом не превышает величину возникновения у сиговых рыб межвидовых гибридов других классов, что косвенным образом подтверждает справедливость заключения о значительном эволюционном обособлении сигa и муксуна, что пока не удалось адекватно подтвердить современными методами генетического анализа. - Слабые оценки генетической дивергенции, отмеченные нами для пары байкальских видов сиговых рыб, подразумевают близкое родство обоих таксонов и соответственно некорректность принятых систематических построений, согласно которым байкальский омуль является подвидом арктического омуля, а байкальский озерный сиг – Подвидом обыкновенного сига. На дендрограмме байкальский омуль и байкальский озерный сиг ассоциируются в отдельный кластер, максимально отстоящий от всех других исследованных нами видов рыб р. Coregonus, в том числе от ветвей, образованных арктическим омулем и обыкновенным сигом (см. рис. 6.2). В последнее десятилетие филогенетическим исследованиям сигоВых рыб Байкала было уделено повышенное внимание. Результаты этих исследований наиболее полно обобщены в работе Д. В. Политова и его коллег (Политов и др., 2010). В целом можно заключить, что полученные нами данные по родственным отношениям байкальского омуля и байкальского озерного сигa и положению обоих таксонов в общей филогенетической схеме сиговых рыб хорошо согласуются с результатами более ранних работ по байкальским видам.
Очевидно, что чир, находящийся в относительно близком родстве с Сигами и муксунами (D-0,105), представляет собой отдельную эволюционную линию сиговых рыб с нижним положением рта. При этом уровень кластеризации для ветви сигов и чиров практически идентичен точке разделения ряпушек и пеляди (D-0,106) – сиговых рыб сконечным положением рта. Все эти виды состоят в более отдаленном родстве с парой байкальских видов сиговых рыб, арктическим омулем, тугуном и нельмой, причем последние три вида в значительной степени генетически дивергировали друг от друга.
Основным отличием приведенной схемы межвидовых взаимоотношений от аналогичных филогенетических построений является положение нельмы относительно видов р. Coregonus. Позиция монотипического р. Stenodus в семействе Coregonidae до сих пор остается предметом дискуссий (см. обзоры Sajdak, Phillips, 1997; Vuorinen et al., 1998). На оснований характерных морфологических признаков (множество зубов, большой рот, максимально приспособленный для хищничества, некоторые потеологические признаки) нелъту традиционно выделяют в отдельный род, альтернативный роду вальков (р. Prosopium) и собственно сигов (р. Coregonus). Тем не менее существует ряд фактов, свидетельствующих о большем сродстве нельмы с сиговыми рыбами р. Coregonus. Известно, например, что р. Stenodus образует естественные гибриды с некоторыми видами р. Coregonus: отмечены гибриды нельмах сиг, нельма хомуль, нельмах муксун (Берг, 1948; Dymond, 1943; Alt, 1971; Reist et al., 1992). Согласно результатам изоферментных анализов, уровни генетической дивергенции между нельмой и видами р. Coregonus (D=0,150 – 0,284, по нашим данным; D =0,305 — по Бодали и др. (1991)) не достигают величин, обычно наблюдаемых при межродовых сопоставлениях рыб (Avise, Aquardo, 1982; Thorp, 1982).
Более того, из включенных в настоящее исследование видов сиговых рыб, помимо наиболее эволюционно обособленного р. Prosopium (уровень кластеризации относительно других видов ~0,6 по результатам наших исследований, ~0,8 – по данным Вуоринена и др. (Vuorinen et al., 1998)), найболее генетически обособленными в семействе оказались представители p. Coregonus – тугун, арктический омуль и байкальские виды сиговых рыб, а не нельма.. В отличие от выводов других авторов (Bodaly et al., 1991; Politov et al., 2004), согласно которым р. Stenodus рассматривается как отдельный, хотя и близкородственный таксон p. Coregonus, наши данные показывают большее сродство двух номинальных родов семейства и возможную искусственность выделения нельмы в отдельный таксон родового уровня. Следует отметить, что сделанный нами вывод подтверждается заКлючениями некоторых других исследователей сиговых рыб (Bernatchez et al., 1991; Reist et al., 1992; Lockwood et al., 1993). На на Среди видов р. Coregonus помимо СИГовых Байкала наиболее генетически обособленными оказались тугун (D-0,250) и арктический омуль (D-0,229), что хорошо согласуется с результатами предыдущих работ по геносистематике сиговых (Ермоленко, 1989а; Bodaly et al., 1991; Бодали и др., 1994). Оба этих вида вместе с исследованными нами ряпушками и пелядью относятся к сиговым рыбам с конечным и верхним положением рта, традиционно объединяемым в подрод Leucichthys. Результаты исследований остеологических признаков были неоднозначны: часть из них подтверждала монофилию сиговых с конечным и верхним положением рта (Shaposhnikova, 1970; Smith, Todd, 1992), другие опровергали эту точку зрения (Norden, 1961).
Данными нашего анализа выделение тугуна, омуля, ряпушек и пеляди в отдельный таксон подродового уровня не подтверждается, поскольку все эти виды не показали признака монофилетической группы — более тесной генетической связи между собой. Выводы о полифилетическом происхождении видов номинального подрода Leucichthys были сделаны и на основании анализов с применением различных методик изучения ДНК (Bernatchez et al., 1991; Lockwood et al., 1993; Sajdak, Phillips, 1997), а также аллозимной изменчивости (Ермоленко, 1989а, б; Bodaly et al., 1991; Бодали и др., 1994; Politoy et al., 2002.2004; Sеndеk, 2002). НапроТив, обнаруженная в настоящем исследовании генетическая близость разны7 видов циповы\о нижним полотенцем рта (подродQoregonus) ?0жет служить доводом в пользу гипотезы о происхождения сига, муксуна и, по-видимому, чира от единого предкового таксона (см. рис. 6.2).
Некоторые противоречия между результатами филогенетических построений, в основу которых положены результаты популяционногенетических исследований, и традиционными взглядами на систематику Сиговых рыб объясняются несовпадением темпов эволюции различных групп признаков, положенных в основы классификаций. Учитывая медленную скорость эволюции проанализированных нами белков-ферментов и значительную морфологическую и экологическую пластичность сиговых рыб даже на внутривидовом уровне, можно предполагать, что во многих случаях генетические данные более точно, чем оценки биологических качеств, отражают реальные родственные взаимоотношения между изучаемыми таксонами (Викторовский, Ермоленко, 1982; Дорофеева, 1989).
Особый интерес представляют внутривидовые исследования сиговых рыб на ареале их обитания. Применение популяционно-генетических подходов к анализу современных популяций позволяет установить движущие эволюционные факторы, которые повлияли на становление отдельных форм, рас, популяций, определить пути расселения видов из разрозненных приледниковых рефугиумов, в которых рыбы переживали периоды оледенения, районы вероятной гибридизации между представителями разных приледниковых рас или номинальных видов (например, между европейской и сибирской ряпушками на р. Печоре), установить историю возникновения симпатрических популяций. Результаты подобных исследований по аллозимной изменчивости сиговых рыб с привлечением наших и ранее опубликованных данных (Перелыгин, 1988; Ermolenko, 1989) для чира, пеляди, сига, европейской и сибирской ряпушек, арктического омуля, тугуна и нельмы были обобщены нами в двух публикациях (Сендек, 2000; Sеndеk, 2002) и впоследствии разработаны более детально для обыкновенного сигa (Sendek, 2004; Сендек, 2005) и ряпушек (Сендек, 2010).