Результаты исследования комбинированной методики кормления (последовательно живым, затем искусственным кормом) и монорационов из живого и различных искусственных кормов
Исследования, направленные на определение возможности кормления нельмы с первых дней жизни живом кормом в сочетании с искусственным, а также монорационами из живого и различных искусственных кормов, были проведены в 2013 г. Биохимические показатели кормов приведены в табл. 21, график проведения экспериментов в табл. 22.
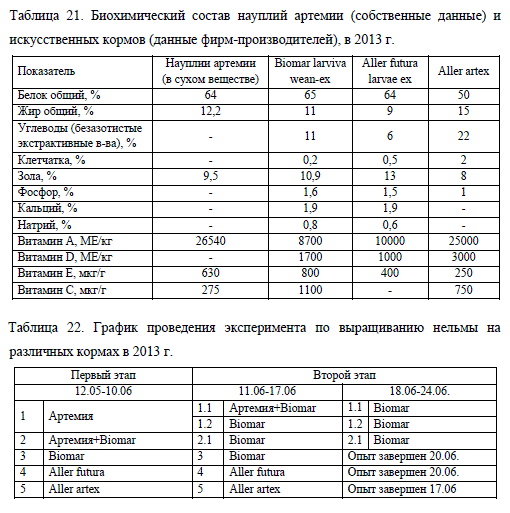
На первом этапе опыта молодь нельмы выращивали в пяти вариантах с применением живых (свежевылупившиеся науплии артемии) и различных сухих искусственных кормов, а также их сочетания. В варианте No1 личинки с первых дней опыта получали живой корм; No2 – сочетание науплий артемии и искусственного корма Biomar larviva wean-ex. В других вариантах использовали только сухие искусственные корма: вариант No3 – Biomar larviva wean-ex; No4 – Aller futura larvae ex; No5 – Aller artex, в состав которого входит экстракт из цист артемии. Продолжительность первого этапа составляла 30 сут., температурный режим в это время находился в диапазоне от 7,7 до 16,7°C, в среднем 12,4°C.
На втором этапе молодь с живого корма (вариант No1) и смешанного кормления (вариант No2) переводили на сухие искусственные корма. Для этого личинок из варианта No1 разделили на две группы: в одной из них (вариант No1.1) в течение первой недели помимо артемии стали добавлять искусственный корм Biomar, а начиная со второй недели – оставили только сухой корм. В другом варианте (No1.2) молодь сразу перевели на искусственный корм. Личинок из варианта No2 также с начала второго этапа стали кормить только сухим кормом (вариант No2.1). Опыт по переводу нельмы с живого на искусственный корм составил 14 сут., температура воды в этот период находилась в диапазоне 16,4-17,4°C при среднем значении 16,7°C. Необходимо отметить, что на втором этапе эксперимента с естественным повышением температуры воды в водоеме и развитием в нем зоопланктона в пищевом комке молоди стали попадаться отдельные экземпляры зоопланктеров, которые составляли не более 1% содержимого желудочно-кишечного тракта нельмы. Исключением явились особи, не перешедшие на питание искусственными кормами, доля зоопланктона в их рационе была существенно выше.
Нельму из вариантов NoNo 3-5 по окончанию первого этапа эксперимента продолжали кормить теми же сухими кормами в течение 10 сут. (до 20.06).
Наполняемость кишечников и избирательность личинками кормов
Наличие корма в кишечниках личинок было отмечено с первых дней эксперимента, а его количество на протяжении первой декады наблюдений во всех опытных вариантах находилось в пределах 0,1-0,2 мг, индексы наполнения при этом колебались от 55 до 950/000 (табл. 23). Следует обратить внимание, что для сиговых рыб в первые дни жизни индекс наполнения кишечников достаточно низок как при питании искусственным (Костюничев, 1986), так и живым кормом (Максимова и др., 1968). В дальнейшем с повышением температуры наполняемость кишечников увеличилась.
На второй декаде опыта при использовании только искусственных кормов индексы наполнения кишечников у личинок были достаточно схожими, за исключением варианта No5, в котором нельма перестала питаться. По итогам третьей декады относительно высокой наполненностью желудочно-кишечного тракта характеризовалась молодь, выращенная на корме Biomar (No3) (табл. 23).
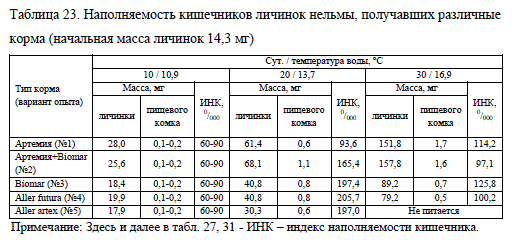
У личинок, имевших в рационе живые корма, масса пищевого комка на второй декаде опыта была выше в варианте, где кормление осуществляли смесью живых кормов с сухими (No2). На завершающем этапе опыта масса пищи в кишечниках личинок из вариантов No1 и 2 была практически равной, что можно объяснить различной массой науплий артемии и гранул искусственного корма.
Еще одной причиной, определяющей массу пищевого комка, является индивидуальная избираемость личинками того или иного корма в варианте No2, что требует отдельного обсуждения. В первые дни нельма, получавшая живой и сухой корм, в подавляющем большинстве питалась только артемией. Лишь у 6% личинок в кишечниках помимо артемии отмечено небольшое количество сухого корма (табл. 24). В последующие 10 сут. эксперимента в варианте с комбинированным кормлением (No2) молодь стала более активно потреблять искусственный корм, его доля в пищевом комке возросла до 45%, а количество потребляющей его молоди – до 76% (табл. 24).
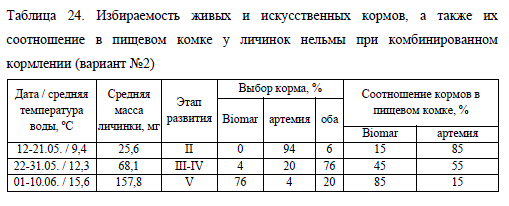
С дальнейшим ростом рыб количество науплий артемии, потребленное нельмой, продолжало сокращаться, а сухого корма, напротив, увеличиваться. По итогам заключительной декады первого этапа эксперимента доля сухого корма в съеденной пище возросла до 85%. Количество личинок перешедших на питание исключительно сухим кормом составило 76%. Темп роста молоди при этом также увеличился и превосходил среднесуточную скорость роста личинок, потребляющих только артемию (табл. 25).
Снижение роли науплий артемии в питании подрощенной (от 100 мг) молоди нельмы может заключаться в небольших размерах рачков в отличие от более крупных гранул искусственного корма, размеры которого увеличивали по мере роста молоди.
Рост, развитие и выживаемость личинок
Рост нельмы на живом корме (вариант No1) и сочетании живого с искусственным (вариант No2) был значительно интенсивнее, чем на одном искусственном. Уже по итогам первой декады суточные приросты личинок, подращиваемых с применением живого корма, составили 5,8-6,7% против 2,3-3,3% в вариантах опыта с использованием только сухих кормов (табл. 25).
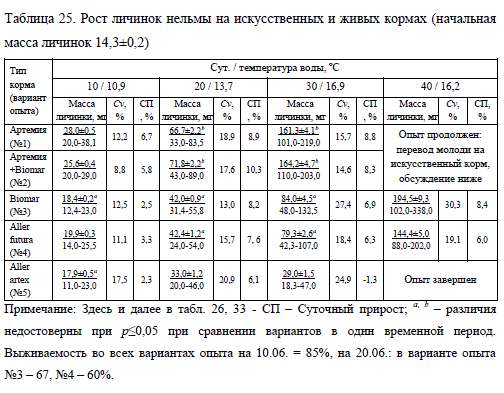
Следует отметить, что использование монорациона из артемии (No1) не выявило заметного преимущества перед комбинированной методикой кормления (No2). Исключением явились первые 10 сут. эксперимента, по итогам которых рост нельмы из варианта No1 опережал по массе молодь из варианта No2 на 8,4%.
В дальнейшем достоверных отличий (при p?0,05) по этому показателю отмечено не было: конечная масса нельмы, выращенной на живом корме (No1) и на сочетании живого и искусственного (No2), была сходной – 161,3 и 164,2 мг, соответственно.
При использовании искусственных кормов Biomar (No3) и Aller futura (No4) масса личинок после 30 сут. выращивания не имела достоверных отличий (при p ? 0,05) и составляла соответственно 84,0 и 79,3 мг (табл. 25). Молодь, получавшая в эксперименте корм Aller artex (No5), уже к середине опыта существенно уступала в росте нельме из других вариантов, а к концу наблюдений и вовсе перестала питаться.
Среди монорационов искусственных кормов по итогам 40 сут. выращивания лучшие результаты были получены в опыте с Biomar: конечная масса молоди на этом корме достигла 194,5 мг против 144,4 мг массы личинок на корме Aller future. Тенденция к более интенсивному росту на корме Biomar проявилась на третьей декаде опыта при средней температуре 15,6oC и существенно усилилась при прогреве воды до 17,0oC (суточный прирост 8,4% против 6,0% на корме Aller future). Вероятно, эти температуры находятся в зоне оптимума для молоди нельмы, в котором в наибольшей степени проявляется эффект от биологически активных компонентов корма (Остроумова, 1988). Именно в этой зоне температур опытная рыба более заметно опережает контрольную по росту.
При температурах ниже оптимальных величин показатели роста рыб в контроле и опыте сближаются, что было отмечено в нашем эксперименте при сравнении искусственных кормов Biomar и Aller Futura.
Причинами, по которым искусственные или живые корма влияют на рост ранней молоди нельмы, по нашему мнению, являются температурный режим и стадия личиночного развития, характеризующая сформированность пищеварительной системы молоди. Так, в первые 10 сут. выращивания при сравнительно низкой температуре воды (9,4°C) максимальный суточный прирост наблюдался у молоди, получавшей только науплии артемии (No1). Этот факт указывает на большее соответствие живого корма биологическим потребностям личинок нельмы, относительно испытываемых искусственных кормов, использование которых не способствует быстрому росту на старте. Это происходит из-за неготовности пищеварительной системы переваривать и усваивать компоненты сухого корма в полной мере, особенно при низких температурах. Возможно, что и сам искусственный корм не вызывает у личинок такого пищеварительного рефлекса, как живые организмы.
На второй и третьей декаде темп роста нельмы на артемии и искусственных кормах был примерно одинаков (табл. 25). Максимальный прирост наблюдался в опыте, где использовалось сочетание сухого и живого корма (No2). Это, по всей вероятности, связано с развитием пищеварительной системы и увеличением ферментативной активности у личинок, что позволяет в большей мере расщеплять и усваивать компоненты корма, особенно искусственного. Кроме этого естественное повышение температуры воды способствует ускорению химических реакций и синтетических процессов в организме, обеспечивающих его рост.
Помимо вышеперечисленных факторов на рост ранней молоди нельмы, безусловно, влияет и биохимический состав кормов. Схожие результаты выращивания на искусственных кормах Biomar larviva wean-ex и Aller futura larvae ex, очевидно, обусловлены приблизительно равным сочетанием в них основных компонентов (табл. 21), в то время как Aller artex существенно отличается от остальных кормов меньшим содержанием белка и бoльшим количеством жира и углеводов.
Биохимический состав науплий артемии по содержанию белка и жира достаточно близок к искусственным кормам Biomar larviva wean-ex и Aller futura larvae ex, однако количество витамина A в них в 2,5-3,0 раза выше, чем в искусственных комах. Функции этого витамина многообразны: он участвует в обмене веществ в организме рыбы, влияет на процессы роста и скелетообразования (Остроумова, 2012). По-видимому, в том числе и по этой причине молодь нельмы, имевшая в рационе живой корм, характеризовалась наиболее быстрым ростом в эксперименте.
По итогам первых 10 сут. выращивания вся нельма в эксперименте перешла на II этап личиночного развития: остаток желточного мешка рассосался, питание стало экзогенным. Дальнейшее развитие личинок было сопряжено со скоростью роста. Так, в конце второй декады в вариантах с использованием науплий артемии были отмечены экземпляры с наполненным газом плавательным пузырем (IV этап личиночного развития). Причем у нельмы, получавшей только артемию, плавательный пузырь был отмечен у 9% особей, а у личинок, имевших в составе пищи сочетание артемии и искусственного корма, – 22%. Остальные личинки в этих группах, как и вся молодь в вариантах с искусственными кормами, имели признаки, соответствующие III этапу личиночного развития: хвостовой плавник принимал трехлопастную форму, в спинном и анальном плавниках появлялись мезенхимные лучи.
В конце третьей декады нельма, выращиваемая с применением живого корма, находилась на завершающем этапе личиночного развития (V этап), характеризующемся появлением в перитонеуме серебристого пигмента гуанина и формированием гомоцеркального хвостового плавника. На искусственных кормах развитие нельмы проходило менее интенсивно, что обусловлено относительно медленным ее ростом. После 30 сут. выращивания в варианте No3 (Biomar) на завершающем личиночном этапе находилось 27% личинок, в то же время на III этапе развития оставалось 23% молоди. При использовании корма Aller Future (No4) развитие нельмы проходило более синхронно: подавляющее большинство молоди (85%) находилось на IV этапе личиночного развития, 15% оставалось на III этапе.
Личинки, получавшие корм Aller Artex (No3), в третьей декаде опыта практически переставали питаться и опускались на дно. Визуально определить причину смертности не удалось. Исследование погибшей молоди под бинокуляром показало отсутствие корма в кишечнике и незаполненный плавательный пузырь. Возможно, компоненты этого корма и их соотношение в нем не отвечали пищевым потребностям личинок нельмы.
О несоответствии, в частности, белкового и липидного состава одного из искусственных комбикормов при выращивании личинок белорыбицы указывала М.В. Михайлова (2001). Автор отмечала, что один из кормов, используемый в их опыте, вызывает изменение в химическом статусе молоди, приводящее к значительному снижению накопления органических и минеральных веществ, что выражается в замедлении роста рыб и их высокой смертности.
Таким образом, корм Aller Artex оказался непригоден для выращивания ранней молоди нельмы. В то же время в литературе есть сведения о положительном опыте использования этого корма на других видах рыб (Piotrovska et al., 2013).
Выживаемость нельмы в эксперименте по прошествии первого месяца была достаточно высокой и составляла около 85% во всех вариантах, за исключением No5. Ранее мы отмечали, что использование сухих кормов увеличивало смертность молоди после наполнения газом плавательного пузыря (IV этап развития, возраст около 30 сут., средняя масса 80 мг) (Лютиков, 2012), однако кормление личинок живыми кормами позволило избежать высокой смертности на данном этапе. В то же время дальнейшее кормление молоди искусственными кормами в вариантах No3 и 4 подтвердило результаты наблюдений прошлых лет – на последней личиночной стадии (возраст 40 сут.) выживаемость в этих вариантах опыта снизилась до 67 и 60% соответственно.
Биологические показатели личинок при их переводе с живого на искусственный корм
При переводе молоди нельмы с живого на полностью искусственный корм заметное преимущество в росте и выживаемости имели личинки, которые с самого начала получали живой корм в сочетании с искусственным (Biomar), а с 11 июня были полностью переведены на Biomar (вариант No2.1) (табл. 26).
Такая молодь превосходила по массе сверстников из других вариантов опыта по итогам первой недели (11-17 июня) в среднем на 26%, а по итогам второй – на 37%. В вариантах No1.1 (поэтапный перевод с живого на искусственный корм) и No1.2 (единовременное замещение живого корма искусственным) достоверных отличий по массе (при p ? 0,05) обнаружено не было. Постепенное исключение из рациона живого корма не отразилось на росте, но сопровождалось более высокой выживаемостью (64%), нежели резкое прекращение его выдачи (53%).
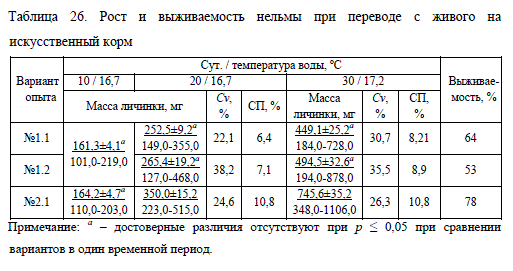
Сокращение артемии в рационе (No1.1) или ее полное исключение (No1.2) привело к снижению скорости роста молоди в среднем на 23%. В дальнейшем темп роста восстанавливался до прежнего уровня, т.е. до момента перевода на искусственные корма, что связано с разреживанием плотности посадки, вызванной повышенной смертностью нельмы. Так, в течение первой половины опыта (с 11 по 17 июня) наблюдалось увеличение смертности с начала недели к ее окончанию: в варианте No1.1 – с 60 до 150 экз./сут., в варианте No1.2 – с 80 до 190 экз./сут., соответственно. Во второй половине опыта элиминация в обоих вариантах заметно снизилась.
Примечательно, что единовременное исключение артемии из рациона (No1.2) приводило к более быстрому росту рыбы, чем при поэтапном сокращении живого корма (No1.1). В то же время не вся нельма начинала активно потреблять искусственный корм после прекращения подачи живого, что приводило к появлению лидеров и отстающих и, как следствие, вело к увеличению вариабельности весовых показателей. Схожая ситуация наблюдалась в варианте No6 при полном переводе молоди на искусственную пищу. Исключение артемии из рациона в варианте опыта с комбинированным кормлением (No2.1) напротив, способствовало увеличению прироста на 23%.
Смертность в течение первой недели перевода на сухой корм снижалась от 35 до 20 экз./сут., на второй неделе смертность была незначительной. Кроме того, вариабельность массы личинок в этом варианте была наименьшей в эксперименте (табл. 26).
Ускорение роста нельмы после прекращения подачи живого корма в варианте No2.1 может говорить о некотором ограничении положительного эффекта от науплий артемии у личинок с возрастом. Мы связываем снижение эффективности использования артемии с ее возможной пищевой неполноценностью (Остроумова, 2014) и мелкими размерами, которые не вызывают пищевого рефлекса у молоди на поздних этапах личиночного развития и требуют большой двигательной активности и затрат энергии. В свою очередь использование искусственных кормов уже на стадии функционирования желудка у личинок нельмы ведет к более быстрому набору массы.
В литературе также встречаются сведения, указывающие на выбор личинками нельмы более крупных организмов. Например, И.И. Смольянов (1956), наблюдавший за развитием ранней молоди нельмы в природе, обратил внимание, что подросшие личинки начинают избегать мелких Cladocera и избирать более крупных Copepoda. Л.С. Богданова (1977), использовавшая при выращивании нельмы в качестве корма Moina macrocopa, также связывала снижение интенсивности питания с небольшими размерами моин. Снижение эффекта от длительного кормления подрощенных личинок белорыбицы зоопланктоном также отмечал М.А. Летичевский (1966). Автор рекомендовал использовать мелкие формы зоопланктона в качестве корма только в первую неделю жизни личинок, а затем переходить на более крупный корм.
Питание нельмы в период ее перевода на сухие искусственные корма зависело от методики перевода. Постепенное исключение артемии из рациона в варианте No1.1 привело к сокращению привычного живого корма в два раза и введениею в рацион искусственного. Нельма в большей степени продолжала поедать науплий, которые составляли до 77% пищевого комка.
В варианте No1.2, где артемию единовременно заменили полностью искусственным сухим кормом, около 20% молоди продолжали питаться исключительно зоопланктоном, поступающим с водой из озера. В их желудках насчитывалось до 80-90 экземпляров представителей ветвистоусых рачков (Cladocera), относящихся к роду Bosmina, что в массе составило всего 1/10 пищевого комка личинок, питающихся искусственным кормом. Масса такой молоди равнялась в среднем 153,7 мг, т.е. была значительно ниже средней, а индекс наполнения кишечника не превышал 530/000. При этом масса перешедшей на питание сухим кормом молоди была 310,1 мг, а индекс наполнения кишечника – 246,80/000 (в табл. 27 представлены средние величины молоди в варианте опыта No1.2).
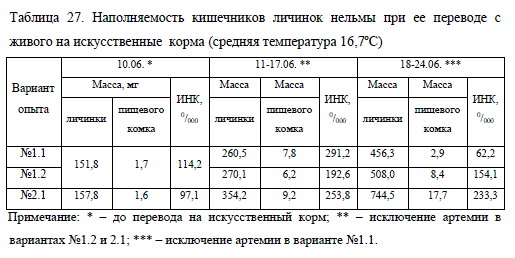
Следует подчеркнуть, что у нельмы из других вариантов в кишечнике также был отмечен зоопланктон, однако его количество относительно пищевого комка в целом было незначительно (менее 1%).
На нежелание личинок сиговых потреблять искусственные корма при их переводе с живого корма указывала Л.М Князева с соавторами (1984). Анализ пищеварительного тракта личинок чира показал, что из смешанного корма они выбирали только живой. После удаления из рациона живого корма поедание искусственных кормов шло неинтенсивно, индекс наполнения кишечника при этом равнялся 9-250/000.
В наших опытах исключение из рациона живого корма в варианте No1.1 на второй неделе эксперимента заметно снизило интенсивность потребления пищи молодью и негативно отразилось на ее росте. Напротив, в варианте No1.2 нельма достаточно быстро адаптировалась к искусственному корму, о чем говорят относительно высокие показатели ее массы и пищевого комка (см. табл. 27). Подобные результаты указывают на необходимость приучения нельмы к искусственному корму на более ранних этапах.
Таким образом, наилучшими результатами характеризовалась молодь, которую изначально выращивали на комбинированном питании, затем полностью переводили на искусственный корм (No2.1). Нельма в этой группе быстро адаптировалась к сухому корму, его содержание в кишечнике в конце опыта было в 3,8 раза больше, чем в варианте No1.1, и в 1,5 раза больше, чем в варианте No1.2 (см. табл. 27). Хорошее потребление корма в варианте No2.1 положительно отразилась на росте и выживаемости молоди, которая превосходила по массе сверстников из других вариантов в среднем на 58% (табл. 26).
Биохимические показатели личинок, выращенных на различных кормах
Важным аспектом в изучении физиологической полноценности молоди рыб является исследование биохимических показателей. Результаты анализа молоди нельмы, выращенной с использованием различных кормов, указывают на однородность ее биохимического состава (табл. 28). Исключением явился вариант No1, в котором только по белку не было достоверных различий (при p ? 0,05) с другими вариантами опыта кроме варианта No3. Также были отмечены различия в содержании жира в теле молоди, выращенной на сухих кормах, ? 3,1% на Biomar (No1) и 2,1% на Aller future (No4), что обусловлено разным содержанием этого компонента в кормах (см. табл. 21).
Сравнение отдельных показателей химического состава устанавливает наибольшее сходство вариантов No1 (артемия) и No4 (Aller future), в них различия обнаруживаются только по содержанию витамина C (табл. 28).
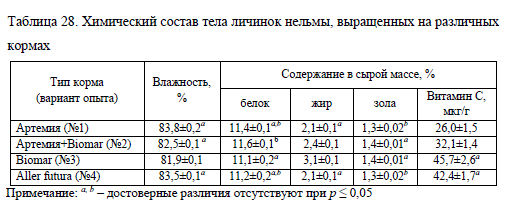
Содержание аскорбиновой кислоты в теле у молоди находилось в прямой зависимости от его уровня в рационе и было достоверно выше в вариантах, в которых применяли только сухой корм. В сухих кормах уровень витамина C был в несколько раз выше, чем в сухом веществе артемии (см. табл. 21). Наименьшее количество этого витамина было отмечено у нельмы, выращенной на живом корме (No1) ? 26,0 мкг/г, тем не менее такое количество находится в пределах нормы. По данным К. Дабровского (цит. по: Остроумова, 2012), дефицит витамина C для личинок сигов C. lavaretus начинает проявляться при снижении его в тканях ниже 20 мкг/г. Молодь, получавшая в рационе как артемию, так и сухой корм (No2), по содержанию витамина C в теле занимала промежуточное значение между вариантами опыта, в которых использовали монорационы из искусственных кормов и науплий артемии (табл. 28). Это объясняется более высоким содержанием витамина в сухом корме, чем в артемии (см. табл. 21). В сухом корме содержание витамина C в 3-4 раза больше, чем в артемии, в то время как у молоди, выращенной на искусственных кормах, количество этого витамина в 1,7 раза больше, чем при использовании живых кормов. Это подтверждают полученные К. Дабровским (цит. по: Остроумова, 2012) сведения о различной динамике накопления витамина C из живых и искусственных кормов. Автор отмечал, что личинки сига в начальный период лучше усваивают витамин С из науплий артемии, чем из искусственного корма. Таким образом, показатели биохимического состава нельмы, выращенной на различных кормах, достаточно однородны, за исключением содержания жира и витамина C в теле.
Подводя итог проведенным исследованиям по выращиванию личинок нельмы на искусственных и живых кормах, а также их сочетании, можно заключить, что использование науплий артемии одновременно с сухими кормами с первого дня питания является наиболее эффективной методикой выращивания. Рыба при таком кормлении характеризуется лучшим ростом и выживаемостью.
Сведения о положительном влиянии сочетания живых и искусственных кормов при выращивании ранней молоди рыб можно найти в литературе. Например, комбинированное кормление белорыбицы искусственным кормом и артемией (живой корм использовался в течение первых 10 сут.) обеспечивало максимальный рост и выживаемость (Михайлова, 2001). Совместное использование коловраток Rotatoria и искусственного лососевого корма в большей степени удовлетворяет пищевым потребностям личинок сига с первых дней жизни до мальковой стадии, чем использование каждого корма в отдельности (Ahmadi et al., 2011).
Комбинированная методика кормления применялась и на других видах рыб. Например, выживаемость личинок судака, которого в течение 20 сут. кормили смесью науплий артемии и кормом РГМ-СС, а последующие 10 сут. только искусственным кормом, равнялась 89% (Михайлова, 1991). Выращивание ранней молоди атлантической трески Gadus morhua на сочетании науплий артемии и сухого корма значительно улучшает рост и выживаемость рыбы и снижает зависимость подрощенной молоди от живой пищи в дальнейшем (Fletcher et al., 2007).
Подобные результаты говорят о благоприятном действии совместного использования живых и искусственных кормов. Такой подход позволяет компенсировать неполноценность каждого корма в отдельности. Более того, дальнейший перевод молоди только на сухие корма происходит лучше при его введении в рацион с первых дней экзогенного питания личинок.