ЭКОЛОГИЧЕСКИЕ АСПЕКТЫ СКАТА ЛИЧИНОК СИГОВЫХ РЫБ ПРИ ДЛИТЕЛЬНОМ МИГРАЦИОННОМ ПУТИ
Покатная миграция личинок (скат) – широко распространенное для рыб явление, в том числе сиговых Coregonidae, считается приспособлени ем к распределению по местам нагула.
Покатная миграция молоди представителей десяти семейств рыб, куда не входили сиговые, подробно рассмотрена Д.С. Павловым и его кол легами (Павлов и др., 1979; 1981, 1985, 2007; и др.). На большом фактическом и литературном материале показана адаптивная природа мигра ций, способствующих расселению вида, увеличе нию площадей нагула и в конечном счете сбалан сированному функционированию популяций в колеблющихся условиях среды. Несмотря на цир кумполярное обитание и важное значение сиго вых рыб как биоресурса, покатная миграция ли чинок сиговых рыб наиболее исследована на при мере лишь нескольких нерестовых рек трех бассейнов (Обский, Байкальский и Анадырс кий), Для североамериканского континента сход ная информация отсутствует. В литературе имеет ся довольно много данных о скате личинок бай кальского омуля (Мишарин, 1953; Краснощеков, 1958; Топорков, 1964; Хохлова, 1965; Шумилов, 1970; Сорокин, Сорокина, 1977; Сорокин и др., 1981; Афанасьев и др., 1981; Щербаков, 1983; Се менченко, Семенчеко,1988) и сиговых р. Анадырь (Юсупов, 1990; Шестаков, 1991а, б). Сведения о скате личинок сиговых рыб в некоторых нересто вых притоках нижней Оби приводятся в работах
В.Г. Иванчинова (1935), В.А. Замятина (1971), П.П. Прасолова (1988), О.А. Госьковой (2010). Более подробно покатная миграция личинок об ских видов сиговых, особенно на нерестилищах, описана авторами статьи (Богданов, 1983, 1987, 2006; Богданов, Богданова, 1984; Богданов и др., 1991; Богданов, Кижеватов, 2000).
Обобщая все имеющиеся в литературе сведе ния, можно сказать, что у полупроходных и ту водных сиговых рыб покатная миграция личинок проходит на различных реках однотипно, что свя зано в первую очередь со сходством биологиче ских аспектов размножения: нерест происходит осенью, а вылупление личинок – ранней весной в период прохождения паводковой волны. Суще ствующие различия ската личинок в разных реках определяются экологическими особенностями динамики освещенности и скорости течения во ды, протяженностью миграционного пути, кор мовыми условиями. При определенных условиях на скате личинки сиговых рыб (омуля, валька) могут переходить на потребление внешней пищи.
Цель настоящей работы – на основе многолет них данных выявить особенности многолетней, сезонной и суточной динамики интенсивности ската, пространственно временной структуры, роста и поведения личинок сиговых рыб при дли тельном миграционном пути.
Исследования покатной миграции личинок сиговых рыб проводили в 1981–2011 гг. в низовье р. Северной Сосьвы (учетный створ находился в 400 км от верхней и 280 км нижней границы нере стилищ), которая считается самым крупным при током нижней Оби, имеет длину 866 км и пло щадь бассейна 89.7 тыс. км2, ее водный режим за висит от выпадения и таяния осадков на Урале (Кеммерих, 1961). Ширина русла в учетном ство ре составляет в паводок 400 м, наибольшая глуби на в межень – 6 м, в паводок – 13.8 м, прозрач ность воды в паводок изменяется от 0.5 до 1.2 м. Ниже учетного створа начинается пойменная си стема нижней Оби – основные места нагула ли чинок (Богданов, 1988).
Пробы брали ежегодно от начала и до конца ската одновременно в разных горизонтах потока и поперечному сечению реки. Всего собрано 3120 проб, определено до вида 58 тыс. личинок. При сборе материала по скату личинок применя ли метод учета дрифта (Пахоруков, 1980; Павлов и др., 1981; Богданов, 1987). Использовали ло вушки типа конусной сети, изготовленной из ка пронового сита No 20, длиной 2.5 м и площадью входного отверстия 0.25 м2. Время экспозиции определяли в зависимости от интенсивности ми грации и засоряемости ловушки от 1 мин до 30 мин (чаще 5–10 мин). Периодичность взятия проб составляла от 1 до 6 раз в сутки. Измерение длины тела личинок проводили на особях, фик сированных 4% ным раствором формалина.
Во время взятия проб регистрировали скорость течения (использовали гидрометрическую вертушку ГР 99), уровень и температуру воды.
Для оценки концентрации покатной молоди в потоке пользовались формулой, предложенной Д.С. Павловым и др. (1981):
М100=m100/Q,
Q = SVt,
М100– количество рыб в 100 м3; m – среднее число рыб в пробах за расчетный период времени; Q – расход воды через сетку (м3/с); S – площадь входного отверстия (м2); V – скорость течения в сетке (м/с); t – время лова (с).
При определении погрешности метода учета численности личинок использовали ошибки, возникающие за счет изменения количества ли чинок в пробах, величины расхода воды и эффек тивности фильтрации ловушки. Общая погреш ность применяемого метода не превышала 40%, чаще была близка к 30% (Богданов, 1987).
кованные рекомендации (Методическое посо бие..., 1974). На анализ взяты личинки пеляди, скатывающиеся в 1981–2001 гг., тугуна, сига пы жьяна и чира – в 1981, 1982, 1985 гг.
Видовую принадлежность личинок устанавли вали по разработанному нами определителю (Богданов, 1998).
Основные нерестилища пеляди Coregonus peled (Gmelin), чира С. nasus (Pallas), сига пыжьяна С. lavaretus pidschian (Gmelin) и тугуна С. tugun (Pallas) Обского бассейна расположены в ураль ских притоках нижней Оби, в предгорных и гор ных участках рек (Москаленко, 1958; Характери стика экосистемы..., 1990). Важнейшие из них на ходятся в р. Северной Сосьве (Богданов, 1998). Личинки с паводковой волной выносятся с нере стилищ, перемещаются в низовье реки и распре деляются по местам нагула. Сравнивая абсолют ную численность покатных личинок во всех ос новных нерестовых притоках нижней Оби (Богданов, 2005), установили, что доля р. Север ной Сосьвы в воспроизводстве обских сиговых рыб составляет 80.0% по пеляди, 45.6% – по чиру, 6.1% – по сигу пыжьяну и 90.4% – по тугуну.
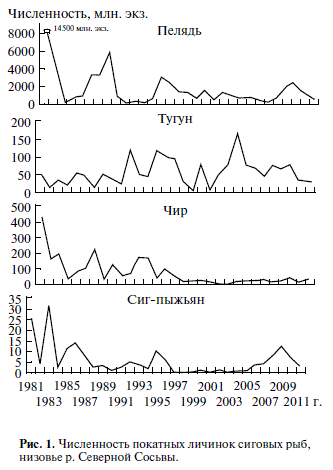
По р. Северной Сосьве скатываются личинки пяти видов сиговых рыб: пеляди, тугуна, чира, си га пыжьяна и нельмы Stenodus leucichthys nelma (Pallas). Среди покатных личинок обычно преобла дает пелядь (средняя многолетняя численность – 1770.1 млн. экз.). Чир, как правило, занимает вто рое место (75.7 млн. экз.). Его доля существенно повышается в годы низкой численности пеляди. Тугун среди покатных личинок стоит на третьем месте (50.5 млн. экз.). Численность личинок сига пыжьяна – самая низкая из сиговых (5.7 млн. экз.), исключая нельму. Личинки нельмы встре чаются единично и крайне редко. Они были зарегистрированы в 1983, 1988, 1989, 1998, 2004 гг. в первые дни покатной миграции.
За 30 лет выявлены четыре пика численности личинок пеляди и каждый последующий был меньше предыдущего (рис. 1). Наблюдается зату хающая амплитуда колебания численности рож даемых поколений. Наиболее значимые повыше ния численности личинок пеляди происходили после годов рекордно высокой водности Оби (по сле 1979 и 2007 гг.). Однако в 2000 е годы ампли туда колебания численности генераций пеляди сильно сократилась за счет повышения числен ности в фазе депрессии и понижения – в фазе подъема (см. рис. 1). У других сиговых рыб анало Эндогенные запасы пищи (наличие и размеры
гичных “волн жизни” (связанных с водностью желтка без жировой капли) определяли у личинок
поймы Оби) не наблюдается (Богданов, Агафо пеляди (1981–2001 гг.). При исследовании экзо
нов, 2001). При общем снижении численности ге генного питания личинок использовали опубли
нераций всех видов полупроходных сиговых рыб
реки; о – дата ледохода.
видов) выносится в русло и далее в пойму Малой Оби и даже достигает дельты Оби (чир), а часть (тугун почти весь) остается в пойме р. Северной Сосьвы (Богданов, 1988). Рис. 1. Численность покатных личинок сиговых рыб, низовье р. Северной Сосьвы.
Скат в низовье реки длится от 10 до 35 сут, чаще – от 16 до 17 сут. В 1980 e годы средняя продолжитель ность ската составляла 18.7 сут, в 1990 е – 16.6 сут, а наиболее сильно снизил численность чир. В р. Северной Сосьве сиг пыжьян после 1980 х го дов стал малочислен, но в последние годы его численность несколько возросла. В отличие от полупроходных сиговых рыб численность личи нок тугуна (туводный вид) устойчиво высокая в последние два десятилетия, отмечены лишь крат ковременные спады.
Личинки, вылупившиеся первыми, скатыва ются медленнее, чем вылупившиеся в пик ската, поскольку в начале паводка скорости течения меньше. Если последним требуется всего 5–6 сут для того, чтобы достигнуть районов нагула, то у первых миграция при возврате холодов затягива ется до 15–20 сут.
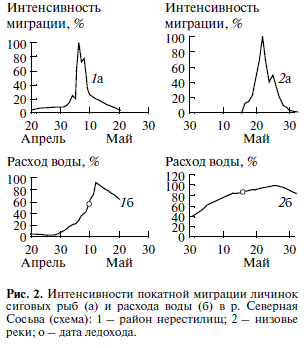
В период ската личинок в низовье р. Северной Сосьвы расход воды был наиболее высоким в 1987 г. (максимум 4800 м3/с) и наиболее низким – в 1984 г. (максимум 1950 м3/с). В районе учетного створа скорость течения воды изменялась за все годы ис следований на стрежне у поверхности от 0.55 до 1.2 м/с, в нижних горизонтах потока – от 0.46 до 1.1 м/с, температура – от 1 до 12°С.
В районе учетного створа личинки появляются после ледохода (рис. 2), через 13–17 сут от начала
1991 в 2000 е уменьшилась до 12 сут. Сокращение про должительности ската связано с устойчивым трендом на снижение численности личинок за многолетний период наблюдений. Первые покат ные личинки появляются обычно 17–23 мая (са мая ранняя дата появления первых личинок – 25 апреля, самая поздняя – 31 мая). Основное их количество скатывается чаще в третью декаду мая, реже – в первую декаду июня. Пик ската все гда происходит за короткий период времени – от 2 до 5 сут. В пик скатывается около 80–90% всех личинок. Среди скатывающихся личинок всегда есть мертвые особи, которые дрейфуют в нижних горизонтах потока. Подсчитано, что в среднем в период покатной миграции гибнет около 1% ли чинок за 10 км пути.
Ежегодно колебания численности покатных личинок происходят независимо от изменений расхода воды в реке (Богданов, 1987). Наиболее интенсивно скат может происходить в различные периоды прохождения паводковой “волны”: при увеличении уровня и расходов воды в реке (чаще всего – 61% от всех лет наблюдений); в период наибольших уровней и расходов воды (24%); в пе риод снижения паводковой “волны” (15%). залития соров полой водой, реже позднее или
Скат личинок проходит круглосуточно, при раньше. Часть личинок (различная у отдельных
этом не отмечено четко выраженного ритма в из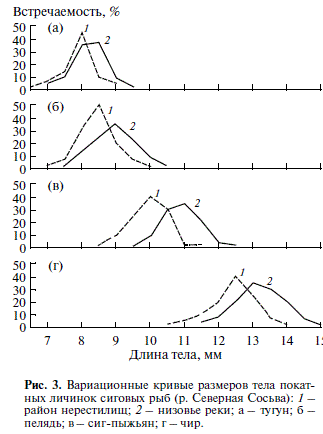
Рис. 3. Вариационные кривые размеров тела покат ных личинок сиговых рыб (р. Северная Сосьва): 1 – район нерестилищ; 2 – низовье реки; а – тугун; б – пелядь; в – сиг пыжьян; г – чир.
менении интенсивности миграции. Наиболее рав номерно распределены по вертикали личинки чи ра и сига пыжьяна. При общем сходстве горизон тального распределения личинок различных видов у каждого из них есть особенности. Тугун распре деляется более равномерно, а пелядь – менее.
Как правило, концентрация покатных личи нок в нижних горизонтах в 0.5–2 раза выше, чем в поверхностных, что прежде всего относится к пе ляди. У чира, пеляди и сига пыжьяна горизон тальное распределение в потоке сохраняется та ким же, как на нерестилищах, – наиболее интен сивный скат происходит на стрежне и у вогнутого берега излучин, тогда как у тугуна распределение в потоке более равномерное. Специфическое рас пределение личинок тугуна в потоке связано с его повышенной подвижностью. Поведение тугуна во время покатной миграции в русле р. Северной Сосьвы направлено на выход из потока, что спо собствует расселению в пойме родной реки, а у пеляди, чира, сига пыжьяна – на удержание в по токе, что приводит к преимущественному рассе лению в пойме Оби (Богданов, 1988, 1992).
Смена поведения личинок в конце миграци онного пути по сравнению со скатом в районе не рестилищ связана с уменьшением объема желточ ного мешка, выполняющего, помимо энергетиче ских, и гидростатическую функцию (Черняев,
Диаметр, мм Доля особей, % 50 40 30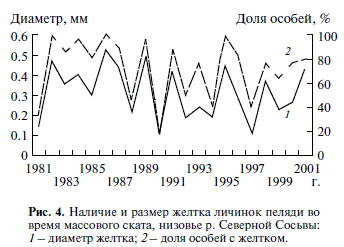
Рис. 4. Наличие и размер желтка личинок пеляди во время массового ската, низовье р. Северной Сосьвы: 1 – диаметр желтка; 2 – доля особей с желтком.
1968), а также с усталостью личинок, которые ста новятся менее подвижными. 20 10 0
7 Средняя длина тела личинок тугуна изменялась в суточных пробах от 7.1 ± 0.08 до 8.5 ± 0.04 мм, пеляди – от 8.0 + 0.03 до 9.9 ± 0.04, сига пыжьяна – от 9.8 ± 0.15 до 11.1 ± 0.13, чира – от 12.2 + 0.03 до 12.9 ± 0.05 мм. Средняя масса особи составляла соответственно 2.6; 3.5; 5.4; 10.7 мг. В низовьях ре ки покатные личинки имеют бо льшую длину те ла, чем в районах нерестилищ (рис. 3). Прирост во время ската от нерестилищ до районов нагула со ставляет у разных видов от 0.3 до 1.0 мм. Личинки, скатывающиеся позднее, имеют в среднем боль шую длину тела. Линейный рост происходит за счет внутренних резервов, в первую очередь за счет желтка. При этом у личинок несколько сни жается масса тела. По размерам и морфологиче ским показателям тела покатные личинки сиго вых рыб в р. Северной Сосьве за все годы были в пределах нормы (Богданов, 1998).
Личинки пеляди, скатываясь по р. Северной Сосьве, к местам нагула подходят каждый год с разным количеством эндогенных запасов, заклю ченных в желтке – депо, из которого личинки черпают энергетические и пластические материа лы (рис. 4). Особи ранних сроков ската имеют их больше, чем особи, скатывающиеся в более позд ние сроки: за 21 год наблюдений в начальный пе риод ската все личинки имели желток в 60% слу чаев (лет), в массовый период – 14%, в последние дни ската – 0%; в разные периоды ежегодного ската количество личинок, сохранивших желток, варьировало в значительных пределах – соответ ственно от 24 до 100%, от 16 до 100% и от 5 до 80%; уровень потери эндогенных запасов от первона чальных изменялся соответственно от 8.6% до 58.1%, от 14.2 до 99.7% и от 99.6 до 99.9%.
Разнокачественность личинок по величине эн догенных запасов при подходе к нагульным водое мам определена неоднородностью таковых при вылуплении (Богданов, 1983; Богданов и др., 1991)
Таблица 1. Коррелятивная связь (p < 0.01) количества эндогенных запасов пищи личинок пеляди массового ска та и абиотических факторов (низовье р. Северной Сосьвы)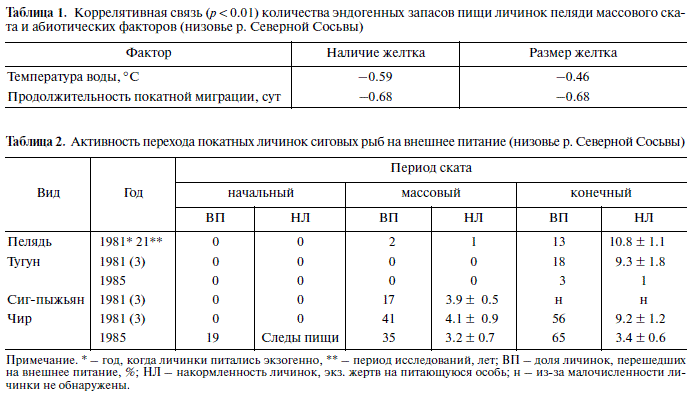
Примечание. * – год, когда личинки питались экзогенно, ** – период исследований, лет; ВП – доля личинок, перешедших на внешнее питание, %; НЛ – накормленность личинок, экз. жертв на питающуюся особь; н – из за малочисленности ли чинки не обнаружены.
и влиянием абиотических факторов (табл. 1). Из
вая величина температуры воды для перехода на представленных данных следует, что, чем выше
внешнее питание для личинок чира и сига пыжья температура воды в реке и чем дольше скатывают
на равна 4–5°С, для пеляди – 8°С. ся личинки пеляди, тем больше они расходуют эндогенной пищи.
Для сохранения жизнеспособности особей при потере эндогенных запасов частично или полностью важен скорейший переход личинок на активное (смешанное или экзогенное) питание. Сиговые выдерживают голодание после полной резорбции желтка при 0.2–1.5°С максимально 10 сут, при 4–5°С – 5 сут (Сергиенко, 1995). Име ются данные (Максимова и др., 1967; Дмитриева, Воинова, 1988; Семенченко, 1988; и др.), что дли тельное голодание (например, у байкальского омуля дольше 15 сут) приводит к необратимым изменениям в организме и гибели личинок, сиго вых рыб даже при наличии пищи. Личинки сиго вых рыб при покатной миграции готовы к приему экзогенной пищи извне за счет функционирова ния ротового аппарата и пищеварительной систе мы, но для этого им нужны благоприятные кормо вые условия (трофический фактор), определяемые в основном температурой воды, концентрацией
На нерестилищах р. Северной Сосьвы личин ки сиговых рыб не переходят на внешнее питание из за экологических условий: температура воды не превышает 2°С; кормовая база отсутствует (Богданова, 2005).
В низовье р. Северной Сосьвы на учетном створе были обнаружены питающиеся личинки всех обследованных видов (табл. 2). Однако ли чинки пеляди и тугуна, которые смогли перейти на потребление экзогенной пищи, отмечены в не большом количестве только в конце ската одного года (1981 г.), отличающегося поздними сроками ската и ранним половодьем. В этот же год во вре мя массового ската питалась небольшая часть ли чинок и сига пыжьяна. За три года наблюдений чир потреблял пищу извне в течение двух лет на протяжении всего ската, активность перехода на смешанное и экзогенное питание потребления пищи была выше, чем у других видов, и возраста ла от начала к концу ската. кормовых организмов и освещенностью. Для ли
Рацион покатных личинок всех видов сиговых чинок сиговых при переходе на внешнее питание
состоял из зоопланктонных (Cyclopoida наупли (смешанное или экзогенное) характерна видоспе
альных и копеподных стадий, половозрелые Acant цифичность по отношению к перечисленным
hocyclops sp., молодь Cladocera – Holopedium gib факторам, что доказано экспериментально в усло
berum, Alonella exisa, Bosmina longirostris, B. longispi виях рыбозаводов (Кугаевская, Сергиенко, 1985;
na) и меропланктонных организмов (larve Chi Широбоков, 1987; Сергиенко, Кугаевская, 1990;
ronomidae и larve Ephemeroptera, с длиной тела ме Сергиенко, 1995). Так, например, нижняя порого
нее 1.1 мм). Первые преобладали в рационе личинок пеляди (98.5%), вторые – остальных ви дов (тугун – 58.1%, сиг пыжьян – 66.8%, чир – 58.9%).
Как видим, в районе исследований наиболее благоприятные условия для перехода на внешнее питание формируются для личинок самого “крупного” вида – чира, что обусловлено прежде всего его способностью переходить на внешний корм при более низких температурах, чем, напри мер, пелядь, и потреблять организмы широкого размерного диапазона с преобладанием в рацио не меропланктона. Стартовым кормом личинок пеляди являются мелкие зоопланктонные орга низмы (не более 0.5 мм) (Норенко и др., 1977; Ко кова, 1978), прежде всего науплиусы веслоногих рачков (Богданова, 1985). За годы исследований численность молоди веслоногих рачков в русле Северной Сосьвы во время ската личинок не пре вышала во время массового ската 0.15 тыс. экз./м3 и 1.00 тыс. экз./м3 – в конце ската, т. е. кормовая база личинок пеляди не успевала развиваться к подходу их с нерестилищ.
Температурные условия за годы наблюдений во время ската личинок не всегда были благоприят ными для питания сиговых, особенно для пеляди: в начале ската вода не прогревалась выше 5°С и в 76% случаев (лет) была ниже 4°С; во время массо вого ската разных лет температура была равна 2.5– 10°С и в 62% случаев (лет) была ниже 6°С; в по следние дни ската вода прогревалась от 6 до 12°С.
Отсюда следует, что в условиях р. Северной Сосьвы наиболее приспособлены к трофическим условиям во время ската личинки тугуна и чира: тугун скатывается раньше других видов при низ кой температуре, поэтому желток резорбируется медленно, при повышении температуры может питаться меропланктонными организмами, нагу ливается “близко” – в пойме родной реки; у чира темп резорбции желтка ниже, чем у пеляди, при мерно в 2 раза (Сергиенко, 1995), при этом он срав нительно активно питается в потоке. Наиболее уяз вимый вид – пелядь. Основываясь на литературных данных (Волкова, 1965; Кугаевская, Сергиенко, 1985; Широбоков, 1987; Семенченко, 1988; Решет ников и др., 1989; Сергиенко, Кугаевская, 1990 и др.), считаем, что отсутствие возможности питаться на скате на фоне значительных потерь эндогенных запасов может привести к повышенной смертности личинок пеляди как при дальнейшей миграции (до 5 сут), так и на местах нагула.
Комплексы причинно следственных связей, реализующих покатные миграции молоди рыб, включают три порядка механизмов (Павлов и др., 2007). Механизмы первого порядка создают пред посылки миграции, второго – реализуют их, третьего – определяют пространственно временную структуру распределения молоди в потоке.
Нами установлено, что у сиговых рыб суще ствуют специфические механизмы первого по рядка, к которым относятся: морфологические адаптации – большой желточный мешок и спо собность личинок к длительной двигательной ак тивности при отсутствии корма; фотореакция и специфическая поведенческая реакция, обеспе чивающая векторизацию движения личинок в потоке. Скат личинок сиговых рыб происходит “пассивно” (механизмы второго порядка), что связано с физической невозможностью сопро тивляться потоку. Однако личинки реагируют на течение и при длительном скате могут в опреде ленной мере изменять свое положение в потоке вплоть до выхода из него (механизмы третьего по рядка). Механизмы, обеспечивающие пассивный скат и корректирующие пространственное рас пределение личинок, у сиговых и других видов рыб во многом сходны.
Скат личинок сиговых рыб в низовье р. Север ной Сосьвы, т.е. при длительном миграционном пути, имеет ряд особенностей, отличающий его от ската на нерестилищах: пик ската происходит по сле ледохода; интенсивность миграции не связана с изменениями расхода воды; нет четкого суточно го ритма; наименьшие концентрации личинок на блюдаются в поверхностном слое потока; видоспе цифическая векторизация движения личинок в транзитном потоке; за счет внутренних резервов происходит линейный рост; эндогенные запасы личинок значительно растрачиваются; личинки сиговых рыб переходят на внешнее питание.
Отметим характерные черты ската на нерести лищах (Богданов, 1983; Богданов и др., 1991; Госькова, 2010): пик ската происходит на сутки – двое раньше ледохода; наблюдается прямая зави симость интенсивности миграции от изменений расходов воды; проявляется суточная динамика в начальный период миграции (при ночных замо розках); преимущественное распределение личи нок в верхних горизонтах потока; векторизация движения личинок по направлению к струям по тока с наибольшими скоростями течения, поло жительная фотореакция; личинки имеют значи тельные запасы эндогенной пищи и не питаются.
Выявлены значительные изменения абсолют ной численности покатных личинок сиговых рыб: пеляди – в 500 раз, чира – в 150 раз, сига пы жьяна и тугуна – в 30 раз. Наиболее устойчивым снижением численности отличается чир, в мень шей степени пелядь. Только у пеляди наблюдают ся четкие “волны” численности при затухающей амплитуде колебаний.
В. Д. Богданов, Е. Н. Богданова Институт экологии растений и животных УрО РАН