К ХАРАКТЕРИСТИКЕ СТРУКТУРЫ ЧЕШУИ И РОСТА МОЛОДИ ХАРИУСОВ НЕКОТОРЫХ СУБАРКТИЧЕСКИХ ПОПУЛЯЦИЙ
В пределах евразийской заполярной зоны встречается два
,вида хариусов- европейский и сибирский (с двумя подвидами).
При значительном ареале их распространения интересно
рассмотреть соответствующее изменение биологических параметров
популяций. Знание жизни рыб на эмбриональных и постэмбриональных
этапах развития является начальной ступенью
'К пониманию биологической продуктивности вида.
Экологические особенности молоди хариусов изучены недо-
статочно. Если эмбриональное развитие и личиночный период
<>нтогенеза освещены подробно (Соин, 1963; Penaz, 1975), мальковый
период- далеко не полно (Мишарин, 1942; Тугарина,
1964, 1967; Тугарина, Топорков, 1967), хотя сведения о разме-
рах сеголеток и годовиков имеются во многих публикациях
(Agaпovic, 1957, 1958; Svetina, 1957; Volk, Vesel, 1961, и др.).
Практически нет данных о формировании чешуйнаго покрова
:и закономерностях роста чешуи молоди хариусов. Лишь Пеньяз
(Penaz, 1975) приводит некоторые материалы по этому воnросу
и фотографии чешуй сеголеток, взятых с хвостового стебля
(очевидно, из-за того, что здесь чешуя образуется раньше), а
не из-под спинного плавника, как принято при изучении хариу
·совых.
Цель настоящего сообщения заключается в установлении
nопуляционной и видовой специфики роста, формирования по·
верхноетной скульптуры чешуи у наиболее северных представи·
телей указанных видов хариуса.
Материал собран 21 июня- 30 августа 1974 г. на реках:
Кожим (среднее течение)- европейский хариус Thymallus thymallus,
Собь (среднее течение)- западносибирский хариус
Thymallus arcticus arcticus и Лена (устье) -восточносибирский
подвид Thymallus arcticus pallasi. Всего исследовано 533 сего.
летка и 142 годовика, для сравнения использовались наши
сборы по восточносибирскому хариусу р. Амгуэмы, осуществленные
в те же сроки в 1973 г. (233 экз.). Отлов производился
мальковым бреднем и сачком. Длина молоди измерялась от
вершины рыла до конца средних лучей хвостового плавника
(у личинок- вся длина). Промеры чешуи и подсчеты склеритов
сделаны при помощи микроскопа МБС-1 при увеличении
Период икрометания у европейских и сибирских хариусов
охватывает почти полгода: с февраля для популяций Северной
Италии по июль для обитателей тундровых озер сибирского
заполярья и некоторых озер Саян (Нойон-холь), но обычно
приурочен к моменту распаления льда с температурным диапазоном
2,5-12,0 0°С (Гундризер, 1967; Зиновьев, 1971, и др.) ..
Инкубационный период при температуре 6-10° С (с повышением
в конце периода до 13-16°) в разных водоемах резко
различается по продолжительности и не всегда находится в
прямой связи с термическим режимом (Миша рин, 1942; Сои н,
1963; Гундризер, 1967, и др.).
Взгляды ученых, изучавших этапы развития хариусов и их
количественный состав, значительно расходятся. Для полевых
исследований, на наш взгляд, вполне пригодна простая схемарасчленения
на этапы, предложенная А. Ф. Коблицкой (1966):
предличинка, раняя личинка, поздняя личинка и малек. Молоди
на первом этапе постэмбрионального развития в наших сборах
не было. Судя по данным П . .Я. Тугариной (1967) для
черного байкальского хариуса, предличинки ( 11-15 мм) малоподвижны
в течение двух суток после выклева, так как находятся
под слоем гальки.
Европейский хариус р. Кожим при длине тела 16 мм вступает
в завершающую фазу этапа ранней личинки. Желточный
мешок полностью исчезает, личинки становятся более подвиж-·
ными, чаще выходят на прогреваемые солнцем песчано-галечные
отмели, где активно питаются. Плавниковая складка еще
большая, дифференцирована на спинной, хвостовой и преанальный
отделы, заметно увеличена в области плавников. Мезенхимные
лучи образуются сначала в хвостовом, затем в спинном
и анальном плавниках. Конец хорды в области хвостового·
плавника изогнут вверх. Грудные плавники хорошо развиты,
брюшные- находятся в зачаточном состоянии. Меланиновая
пигментация имеется в основном на дорсальной стороне тела,
меньше ее на боках. Сгущение меланофоров образует над головным
мозгом округлую темную «шапочку».
В дальнейшем происходят значительные морфафизиологические
преобразования личинок. Форма тела личинок постепенно
приближается к форме тела взрослых хариусов, позволяющей
активнее передвигаться и кормиться. Плавниковая складка
вначале исчезает между спинным и жировым плавником
(20 мм), затем перед спинным (25 мм) и после этого- перед.
брюшными плавниками (30 мм). Преанальная складка исчезает
последней при длине тела 35-40 мм. В это же время окон-
чательно формируются брюшные плавники и в хвостовом плав.
нике образуются две лопасти. Мезенхимные лучи всех плавников
превращаются в костные. При длине около 35 .м.м на боках
тела становятся хорошо выраженными 9-11 поперечно вытянутых
пятен, характерных и для пестряток лососей. В области
боковой линии появляется чешуя. Личинки превращаются в
мальков. Мальки трех изученных популяций, относящихся к двум
видам хариусов ·с двумя подвидами, похожи по форме тела,
окраске, распределению пигментных пятен, величине и форме
nлавников, хотя у взрослых рыб последние показатели различаются.
Вместе с тем у мальков европейского хариуса меньше
величина спинного и парных плавников и короче верхняя
челюсть, не заходящая за передний край зрачка, у мальков
западносибирского хариуса она достигает середины зрачка, и
у восточносибирского доходит до заднего края зрачка.
У части мальков европейского хариуса из р. Кожим чешуя
nоявляется, когда длина тела достигает 31 .м.м, хотя для центральнаевропейских
популяций констатировано более раннее ее
появление- при длине 27,5 .м.м (Репаz, 1975). Первая чешуя,
nредставленная базальными пластинками, располагается вдоль
боковой линии на хвостовом стебле. При длине 35 .м.м боковая
линия полностью покрывается первичными чешуйными пластинками
и образуется первый ряд чешуи над боковой линией на
хвостовом стебле. При длине тела 40-41 с.м на хвостовом стеб.
ле кожимского хариуса имеется по два ряда базальных пластиLНОК
над и под боковой линией. У западносибирского и восточно
·Сибирского хариусов (реки Собь и нижняя Лена) чешуя появ.
ляется позднее, чем у европейского -при длине 35-38 .м.м, но
.затем формируется быстрее, поэтому у мальков длиной 41 .м.м
nлощадь тела, покрытая чешуей, больше. В дальнейшем нарастание
чешуи происходит в краниальном и дораовентральном
!Направлениях (Penaz, 1975). В последнюю очередь чешуя
локрывает затылочную и грудную части тела.
Формирование склеритов на разных участках чешуи и тела
nроисходит неравномерно. При длине тела 42 .м.м на чешуе в
начале спинного плавника в первом ряду над боковой линией
имеются одна базальная пластинка и один склерит. В том же
месте, но во втором ряду над боковой линией, обнаруживается
лишь одна базальная пластинка. У заднего края спинного плавника
в первом ряду на чешуе имеется, помимо базальной пластинки,
два склерита. На чешуе хвостового стебля из первого
ряда прирост составляет три склерита. Хвостовой стебель поляостью
покрывается чешуей при длине тела 47-50 .м.м. Над
грудным плавником в это время наблюдается только два ряда
<чешуи над и под боковой линией.
Возможно, для более точного определения возраста и рас<
числения темпа роста необходимо знать не только характер
10бразования чешуи при достижении мальком определенного
Таблица 1
Размеры и количество склеритов на разных участках тела
сеголеток хариуса р. Соби, .м.м

• В числителе- минимальный диаметр, в знаменателе- максимальный.
размера, но и закономерности формирования годовых зон, осо-·
бенности склеритообразования в разных секторах чешуи особенно
на первом году жизни.
По характеру формирования чешуйнога покрова и образования
склеритов на разных участках тела западносибирский
(р. Собь) и восточносибирский хариусы (реки нижняя Лена и
Амгуэма) похожи на европейского хариуса р. Кожим. Размеры
чешуи во втором ряду над боковой линией несколько больше.
чем в первом, несмотря на меньшее число склеритов и соответственно
большую величину базальной пластинки (табл. 1).
Количество образующихся склеритов непосредственно коррелирует
с ростом рыбы. У западносибирского хариуса из
р. Соби с увеличением длины тела в первый год жизни в среднем
на 10 мм на чешуе образуется один склерит, а у европейского
хариуса южных популяций ареала- примерно в два-три
раза больше. В среднем у сеголеток хариусов Заполярья за
месяц «период открытой воды» формируется два склерита.
Нарастание склеритов по четырем разным секторам чешуи
происходит неравномерно. Сначала они формируются в боковой
области чешуи, затем в передней и в последнюю очередь в.
задней. В передней части чешуи склериты вдвое уже, чем в каудальной.
Годовая зона северных популяций хариуса чаще всегО>
Таблица 2
Зависимость размеров чешуи и баэа.лыiоЙ плаСтltикн (мм) or дЛИны сеrоле'rок хариуса р. С~и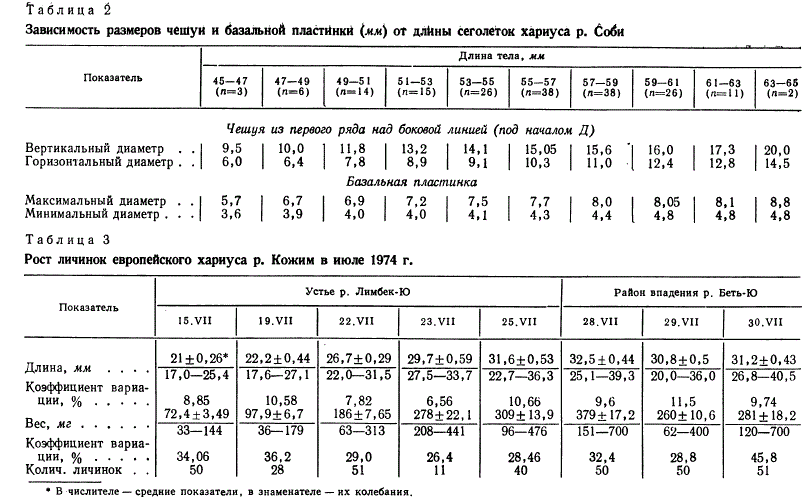
• В ·числителе- средние покаэатели, в знаменателе- их колебании.
Таблица 4
Рост сеголеток некоторых популяций эападно- и восточносибирского хариуса-
Р. Собь Р. нижняя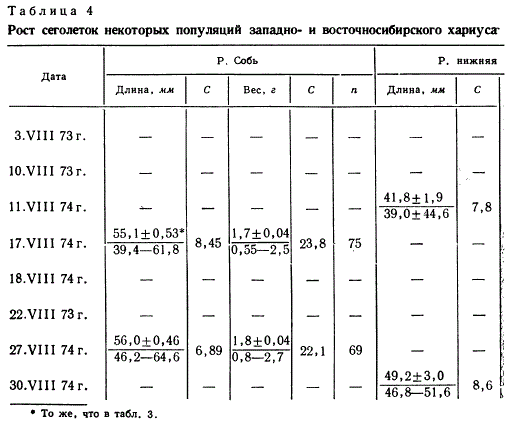
• То же, что в табл. 3.
завершается двумя суженными склеритами, которые могут не·
образовывать полного кольца и выклиниваться при переходе
с переднего сектора на боковой. Подобная форма годовой зоны
бывает у сигов р. Нижнего Енисея (Нейман, 1959, 1961), озерКольского
п-ва (Решетников, 1966), р. Амура (Подушка, 1970)
и у восточносибирского хариуса р. Амгуэмы.
Нередко на чешуях обнаруживаются разрушенные, прерывистые
склериты (особенно по переднему и боковому секторам)_
Это свидетельствует о неблагаприятных условиях обитания
молоди в отдельные периоды развития. Такое же явление отмечала
Л. Ф. IUентякова (1966) при пересадке рыб из аквариумов
:в пруды или обратно при резком изменении температурных
и кормовых условий.
Размеры чешуи и базальной пластинки равномерно увеличиваются
с ростом тела (табл. 2). Вместе с тем величина базальной
пластинки у годовиков практически одинакова, несмотря
на большие различия в их длине. Иногда внутри базальной
пластинки годовиков (и старших по возрасту рыб) видно полуразрушенное
небольшое кольцо, диаметр которого соответствует
размеру базальной пластинки у медленно растуrцих сеголеток.
Поэтому можно предположить, что базальная пластинка
сохраняется только на ранних стадиях развития чешуи, а к
концу первого года жизни разрушается.
Годовая зона на чешуе хариусов из рек Кожим, Собь и
Лена (нижнее течение) формируется в июле, когда длина тела
рыбы 65-70 мм. Мальковая зона отсутствует; в первой годовой
зоне насчитывается три- восемь склеритов, размеры зоны по
переднему сектору колеблются от пяти до девяти делений окулярмикрометра.
При обратном расчислении роста методом
Е. Леа обнаруживаем, что эта зона соответствует длине годовиков
26-65 мм. Полученный первый размерный интервал
может отрицать идентификацию этой зоны как годовой, поскольку
при таких размерах чешуя еще полностью не сформирована.
Следовательно, метод обратных расчислений несовершенный.
Возможно, что отсутствие малькового кольца на чешуе является
специфичной чертой наиболее северных популяций хариуса.
Смена характера питания или местообитания на чешуе малька
не отражается в связи с укороченностЪю периода роста рыбы
и чешуи и небольшим количеством склеритов (вдвое- втрое
меньше, чем у южных популяций и у популяций с оптимальными
условиями в первый год жизни).
В условиях наиболее низких летних температур появляется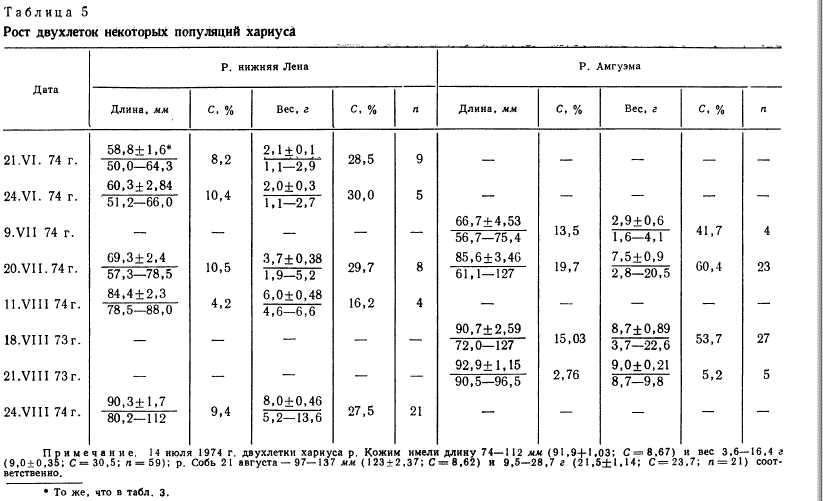
значительная часть молоди, зимующая без чешуйнаго покрова_
Так, среди молоди, выловленной в притоках р. Амгуэмы в конце
вегетационного периода ( 19-22 августа 1973 г.), 35-58%
составляли особи длиной менее 35 мм (иногда 25,2-43,5 мм).
Лишь некоторые рыбы имели в это время неполный чешуйный
покров. У многих двухлеток, отловленных в конце августа в.
р. Амгуэме, не было зарегистрировано первой годовой зоны_
Лишь осенью проявляются суженные склериты.
При изучении роста молоди всех исследованных популяций
обращает на себя внимание значительная индивидуальная изменчивость
показателей, начиная уже с личиночного периода
(табл. 3 и 4). Так, у личинок европейского хариуса р. Кожим
коэффициент вариации (С) длины тела составляет от 6,56 до·
11,5, а веса тела от 26,4 до 45,8. Коэффициенты вариации у
сеголеток западносибирского хариуса р. Соби в августе несколько
меньше- от 6,89 до 8,45 (длина) и от 22,1 до 23,8 (вес)_
Вариабельность длины тела и веса личинок хариуса р. Амгуэмы
приблизительно такая же, как и у кожимского хариуса. Такие
же или большие отклонения в росте отмечены П. Я. Тугариной
( 1964, 1967) для молоди хариуса Иркутского водохранилища и
южных притоков оз. Байкала.
Внутрипопуляционная дифференциация по темпу роста приводит
в дальнейшем к значительной вариабельности всех
остальных признаков популяции, позволяя шире использовать.
экологическую нишу. Средние значения роста сеголеток минимальны
в р. Амгуэме, несколько больше в р. нижней Лене и
заметно выше у европейского хариуса р. Кожим и у западносибирского-
в р. Собь. Замедленным ростом отличаются и
хариусч1 большинства притоков оз. Байкала и Иркутского водохранилища
(Тугарина, 1967). Высокие скорости роста отмечаются
у молоди центральнаевропейских популяций хариуса
(Penaz, 1975, и др.). Тенденции роста, показанные на примере
сеголеток, сохраняются и на втором году жизни (табл. 5).
Очевидно, в целом сибирские популяции хариуса характеризуются
более медленным ростом.
Выводы
1. Видаспецифичность в структуре чешуи у молоди (до
1~года) европейского, западно- и восточносибирского хариусов
не обнаруживается. Наиболее северные популяции отличаются
меньшим количеством склеритов на чешуе. Часть молоди зимует
без чешуйнаго покрова. Мальковые кольца на чешуе хариусов
этих популяций отсутствуют.
2. Морфологически сеголетки двух видов и двух подвидов
хариуса различаются лишь по небольтому числу признаков.
Для европейского хариуса характерны меньшие размеры плавников,
короткая верхняя челюсть и более конечный рот. Маль-
ковый период во всех исследованных популяциях начинается
тогда, когда длина тела хариуса становится 33-37 мм.
Он приходится на разные сроки- от конца июля (р. Кожим)
до конца августа (реки нижняя Лена и Амгуэма).
3. Формирование чешуйнога покрова происходит в краниальном,
дорзальном и вентральном направлениях от боковой линии
в районе хвостового стебля.
4. Молодь хариусов наиболее северных местообитаний меньше
по длине и весу в первое и второе лето жизни в отличие от
центральнаевропейских и южных популяций ареала. Наиболее
медленнорастущие популяции восточносибирского хариуса.
5. Внутрипопуляционная изменчивость длины и веса тела
на первых этапах онтогенеза значительна.