Влияние разных плотностей посадки и режимов кормления на биологические показатели нельмы
Помимо изучения влияния внешних факторов среды на молодь рыб в ранний период постэмбрионального развития, необходимо при их выращивании в индустриальных условиях исследовать такие важные элементы биотехники, как плотность посадки и режим кормления. Действие плотности посадки на особь проявляется непосредственно на уровне тактильных, химических и других контактов и опосредованно – например, в истощении кормовой базы (Гершанович, Тауфик, 1991). Завышенная плотность посадки действует на рыб угнетающе, а низкая ведет к снижению продуктивности выращивания.
Определение суточной нормы корма имеет не меньшее значение, чем количество рыбы в лотке. Избыточное кормление приводит к непроизводительным затратам, загрязнению емкостей и повышению себестоимости выращиваемой рыбы, а недостаточное – к неполной реализации потенциала ее роста. Более того, с помощью правильно подобранной суточной нормы корма можно повысить интенсивность выращивания молоди и улучшить ее физиологическое состояние (Шумилина, 1986). В связи с этим нами была поставлена цель – определить оптимальные для роста и выживаемости плотность посадки и суточную норму корма при выращивании личинок кубенской нельмы в лотках.
Плотность посадки. Для определения оптимальной плотности посадки личинок содержали в лотках размером 4,2?0,7?0,7 м, уровень воды в них устанавливали 0,4 м. На момент начала опыта вся молодь перешла на питание внешним кормом, ее возраст равнялся 10 сут., масса 12,9 мг. С первых дней экзогенного питания и во время эксперимента личинки получали искусственный экструдированный корм Larviva фирмы Biomar в виде микрогранул. Опыты проводили в трех вариантах: 25 тыс. экз./лоток (21,3 тыс. экз./м3), 30 (25,5) и 35 тыс. экз./лоток (29,8 тыс. экз./м3). По мере роста личинок проводили разреживание плотности, поводом для этого служило снижение кислорода на вытоке в лотке с максимальным количеством молоди до 7,0 мг/л, что является близким значением к допустимой минимальной концентрации кислорода в воде для сиговых рыб (Костюничев и др., 2012). Таким образом, спустя 20 сут. после начала эксперимента плотность посадки во всех вариантах опыта была сокращена в два раза, т.е. с 25 тыс. экз./лоток до 12 (10,2 тыс. экз./м3); с 30 – до 15 (12,8); с 35 – до 18 (15,3 тыс. экз./м3). Длительность эксперимента составляла 52 сут.
Для определения названия вариантов опытов в дальнейшем мы использовали показатели плотности посадки или количества корма, соответствующих их настоящему значению в определенном варианте. Например, плотность посадки, равная 15 тыс. экз./лоток соответствует варианту опыта No15.
Результаты выращивания личинок нельмы при различной плотности посадки представлены в два этапа: при начальной плотности (17.05.-06.06.) и после ее разреживания (07.06.-07.07.). На первом этапе при количестве рыбы в лотке, равной 25 и 30 тыс. экз., суточные приросты и показатели массы молоди были достоверно выше (при p ? 0,05), чем в опыте с максимальной плотностью (табл. 16). Скорость роста отражалась на личиночном развитии нельмы: в вариантах опыта No 25 и 30 на стадии наполнения плавательного пузыря газом (IV этап) находились 24 и 30% молоди, соответственно. В лотке с плотностью посадки 35 тыс. экз. на этот этап перешли 13% особей. Остальная молодь в эксперименте оставалась на III этапе личиночного развития.
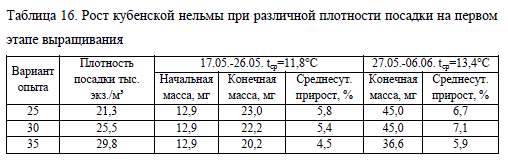
После разреживания посадочных плотностей сохранялась тенденция к более быстрому росту молоди в лотках с меньшим количеством рыбы (табл. 17). Однако в начале второго этапа наблюдений среднесуточный прирост и масса молоди при низкой плотности посадки (12 тыс. экз./лоток) была наименьшей. По-видимому, это можно объяснить чрезмерной разреженностью личинок в лотке, что существенно сокращает вероятность встречаемости нельмы с кормом.
С увеличением биомассы темп роста молоди в этом варианте опыта повысился и к окончанию наблюдений имел наибольшие значения в эксперименте. При плотности посадки 15 тыс. экз./лоток пик роста был достигнут на втором отрезке этапа, однако в итоге масса молоди занимала промежуточное значение между двумя другими вариантами, а показатель среднесуточного прироста был наименьшим. Что связано с относительно быстрым ростом личинок и увеличением ихтиомассы в лотке выше оптимальных значений, что привело к ухудшению условий содержания и угнетению роста рыбы.
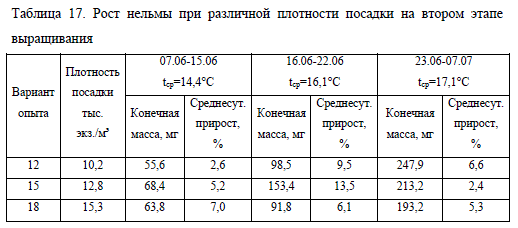
Выращивание нельмы при высокой плотности посадки (18 тыс. экз./лоток) отрицательно сказывалось на ее скорости роста, несмотря на высокие значения этого показателя в первую неделю после разреживания плотности. В дальнейшем показатели среднесуточного прироста снижались, что негативно отражалось на индивидуальной массе личинок, которая по итогам эксперимента была наименьшей в опыте и равнялась 193,2 мг (табл. 17).
Развитие и выживаемость молоди в опыте также зависели от скорости роста и количества рыбы в лотке. На заключительном отрезке наблюдений при минимальной плотности посадки (вариант No12) на мальковой стадии находилось 72% молоди, выживаемость при этом составила 75% (за весь период эксперимента). В варианте No15 на стадию малька перешло 63% молоди при выживаемости 70%, в варианте No18 – 22%, процент выжившей молоди составил 60.
Низкая выживаемость и невысокий темп роста при максимальной плотности посадки (вариант No35) отразились на рыбопродуктивности, которая на первом и втором этапах выращивания имела наименьшие значения в этой группе (табл. 18).
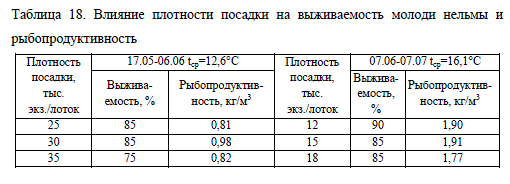
Учитывая показатели выживаемости, роста и рыбопродукции при выращивании ранней молоди нельмы в индустриальных условиях с использованием экструдированных кормов, лучшие результаты были получены при начальной плотности посадки 30 тыс. экз./лоток (25,5 тыс. экз./м3) с последующим сокращением плотности до 12 тыс. (10,2 тыс. экз./м3). Разреживание численности нельмы проводили на 20 сут. эксперимента при индивидуальной массе молоди 45 мг.
Режим кормления. При определении суточного рациона корма использовали личинок в возрасте 20 сут. средней массой 21,0 мг. Их развитие соответствовало III этапу личиночной стадии – дифференциация плавниковой каймы, появление в плавниках мезенхимных лучей и формирование трехлопастного хвостового плавника. Опыт включал в себя четыре варианта, в которых личинки получали корм в количестве 7%, 9,5, 12 и 15% от массы тела. Нельму содержали в лотках размером 215?40?17 см при плотности посадки 3 тыс. экз./лоток (23,3 тыс. экз./м2).
Продолжительность опыта равнялась 40 сут. Температура воды в период наблюдений изменялась в диапазоне 9,7-18,6°C (в среднем – 14,7), содержание кислорода – 8,3-9,6 мг/л (в среднем 9,0). Результаты определения оптимальной суточной нормы корма для личинок нельмы приводятся поэтапно. На первом этапе выращивания (27.05.-09.06.) максимальная норма кормления (вариант No15) обеспечивала наиболее быстрый рост личинок, но отрицательно сказалась на их выживаемости (табл. 19). На втором этапе (10-21.06.), напротив, среднесуточный прирост в этом варианте опыта был самым низким, что связано с увеличением ихтиомассы за первый период выращивания и ухудшением газового режима в связи с избыточным кормлением.
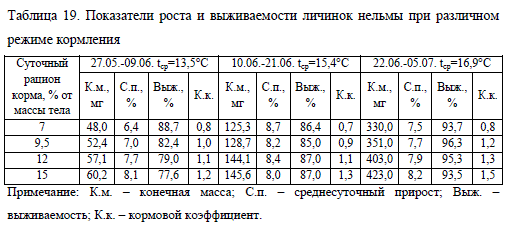
По нашим наблюдениям молодь нельмы не подбирает гранулы со дна. Излишек корма, скапливающийся на дне лотка, не только потребляет кислород в процессе окисления, но и является субстратом для развития патогенной микрофлоры, ослизняющей жабры рыбы (Богданова, 1981). В других вариантах в этот период мы наблюдали увеличение скорости роста, что обусловлено началом функционирования желудка у личинок.
На заключительном этапе выращивания (22.06.-05.07.) по мере развития молоди и повышения ее резистентности к неблагоприятным факторам среды происходило увеличение темпа роста и снижение смертности. Исключением явился вариант No7, где был отмечен самый низкий показатель среднесуточного прироста и выживаемости, что указывает на недостаток корма. Лучший темп роста установлен в варианте No15 (избыточное кормление), что связно с сокращением плотности посадки из-за повышенной смертности молоди в предыдущие периоды. Сокращение количества личинок в лотке обеспечивало более благоприятные условия для их последующего развития. Подобный эффект для нельмы был отмечен нами ранее (Лютиков, 2012). Относительно высокие значения суточного прироста и тенденция к их увеличению в варианте No12 могут служить показателем нормального режима кормления, соответствующего пищевым потребностям личинок нельмы с момента их перехода на питание внешним кормом до мальковой стадии (при средней температуре воды 15°C).
Обобщенные результаты опыта, представленные в табл. 20, подтверждают наши выводы. Рыбопродуктивность, полученная с использованием суточной нормы корма равной 12%, незначительно уступает этой величине в опыте с избыточным кормлением (No15). При этом количество корма, израсходованное за время эксперимента в опыте No12, меньше на 16% по сравнению с опытом No15, что привело к уменьшению кормового коэффициента.
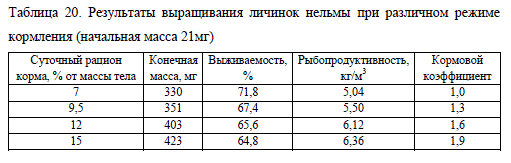
В вариантах опыта с относительно низкими суточными нормами кормления (No7 и No9,5) наблюдалось отставание молоди в росте и снижение рыбопродуктивности. Однако сокращение рациона повысило выживаемость, что, вероятно, связано с благоприятным состоянием гидрохимического режима, обусловленного достаточно полным освоением корма личинками. Таким образом, при анализе результатов выживаемости, рыбопродукции и кормовых затрат при выращивании ранней молоди нельмы в индустриальных условиях от 20 до 400 мг с использованием экструдированных кормов, нами был определен оптимальный суточный рацион равный 12% от массы рыб (в температурном диапазоне 13-17°C).
На основании исследований, проведенных на ранней молоди кубенской нельмы, впервые дано описание ее личиночного развития при выращивании в индустриальных условиях на искусственных кормах. Изучено воздействие солнечной инсоляции на рост, развитие и выживаемость молоди нельмы. Полученные результаты позволили сделать вывод, что солнечная радиация оказывает пагубное влияние на личинок раннего развития, из-за отсутствия у них в теле гуанинового пигмента, который появляется позже и обеспечивает защиту от инсоляции. Кроме того, поведенческие реакции личинок не способствуют снижению негативного эффекта освещенности – личинки не укрываются в затененных участках лотка, а свободно распределяются по всей его площади.
Эксперименты, направленные на исследование биологических показателей нельмы, выращенной под влиянием различных плотностей посадки и количества скармливаемого корма, позволили определить оптимальные значения этих параметров при выращивании личинок нельмы на искусственных кормах. Наиболее предпочтительная плотность посадки для личинок нельмы в начале выращивания составила 30 тыс. экз./лоток (25,5 тыс. экз./м3), по достижении молодью массы 45 мг (20 сут.) оптимальной плотностью является 12 тыс. (10,2 тыс. экз./м3). Суточное нормирование корма, давшее лучшие результаты при подращивании личинок нельмы от 20 до 400 мг, равнялась 12% от массы рыб (в температурном диапазоне 13-17°C).