Влияние температурного режима на эмбриональное развитие нельмы
Влияние температуры на развивающийся организм общеизвестно – повышение температуры (в определенном диапазоне) ускоряет темп эмбриогенеза рыб, понижение, напротив, замедляет этот процесс. Сиговые в этом отношении не являются исключением, скорость их эмбрионального развития также сопряжена с температурой окружающей среды (Борисов, Крыжановский, 1955; Вернидуб, 1956; Яндовская, Тихонова, 1961; Черняев, 1968; Городилов, 1969; Brooke, 1975; Буланов, 1976; Мешков, Лебедева, 1977; Игнатьева, 1979; Решетников и др., 1989; Костюничев, 1997; Богданов, 2006).
Развитие икры сиговых рыб может проходить в достаточно широком диапазоне температур (Лебедева, 1971; 1983), однако зона температурного оптимума существенно уже. Так, Дж.В. Прайсом (Price, 1940) при инкубации икры сельдевидного сига Coregonus clupeaformis в температурных условиях от 0 до 12°C было показано, что успешное развитие этого вида происходит от 0,5 до 6°C. Оптимальные температурные условия развития икры сельдевидного сига были определены Л.Т. Бруком (Brooke, 1975) и соответствуют диапазону температур 3,2-8,1°C. Оптимальные температуры развития икры пеляди лежат в диапазоне 2-5°C, верхний порог развития составляют 7-8°C (Лебедева, 1974, 1985).
Под оптимумом понимают количественное выражение фактора (например градусо-дни), соответствующее потребностям организма и обеспечивающее наиболее благоприятные условия для его жизнедеятельности (Шилов, 2000; Вербицкий, 2008). Температурный оптимум для рыб видоспецифичен и определен экологическими особенностями обитания вида в естественной среде.
Определение температурного оптимума для развивающегося организма может служить обоснованием к его успешному воспроизводству в искусственных условиях. Как правило, в литературе приводятся сведения по изучению эмбрионального развития рыб при статичных температурах, в то время как в природе температурный режим достаточно динамичен. В настоящее время накоплен богатый экспериментальный материал, свидетельствующий о том, что наилучшим образом жизнедеятельность организмов протекает при некотором изменении параметров среды в пределах толерантного диапазона, а не при поддержании постоянных условий с заданными оптимальными значениями (Константинов, 1997; Вербицкий, 2008; Лукиянов, 2010).
Работы, направленные на изучение влияния астатичности температурного фактора, проводились, главным образом, на молоди рыб (Константинов, Шолохов, 1993), в то время как исследованию влияния этого фактора на эмбриональное развитие рыб было уделено заметно меньше внимания. В связи с этим изучение воздействия температурного режима на инкубацию икры при искусственном воспроизводстве рыб носит актуальный характер. Исследование эмбрионального развития кубенской нельмы в различных температурных режимах были проведены нами в период с 2010 по 2014 гг. При анализе полученных результатов также были использованы данные Д.П. Буланова (1976). Средние температуры воды в период исследований представлены в табл. 5.
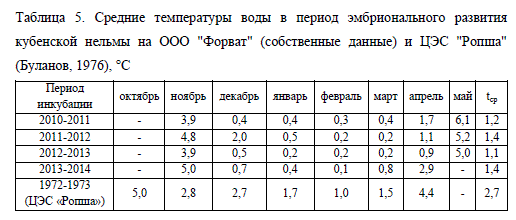
Результаты исследований показали, что эмбрионы, получавшие схожее количество тепла, выраженного в градусо-днях, находились на различных этапах эмбрионального развития. Возраст эмбрионов в эти периоды также мог существенно отличаться (табл. 6). Однако на более ранних этапах эмбриогенеза (до окончания гаструляции) развитие в различных температурных режимах проходило достаточно синхронно – замыкание "желточной пробки" наблюдалось на 11-13-е сут. при 59-74 градусо-днях (табл. 6).
По мере развития икры температура все больше отражалась на скорости эмбриогенеза. Так, в 2012 г., когда был отмечен наиболее холодный температурный режим, отставание на различных этапах эмбрионального развития в сравнении с 2011 и 2013 гг. доходило до 15-19 сут. (формирование жаберных лепестков), а количество полученного тепла было меньше на 21-51 градосо-дней (табл. 6). Темп развития эмбрионов нельмы в 2010 г. был схож с таковым в 2012 г. вплоть до начала тока крови по жаберным дугам, однако относительно высокие температуры воды на последующих этапах эмбриогенеза (см. табл. 5) определили более быстрое развитие зародышей в период 2010-2011 гг. Несмотря на это, ток крови по жаберным лепесткам во все годы наблюдений был отмечен в схожем возрасте, но сумма полученного тепла при этом существенно варьировала – от 148 до 215 градусо-дней.
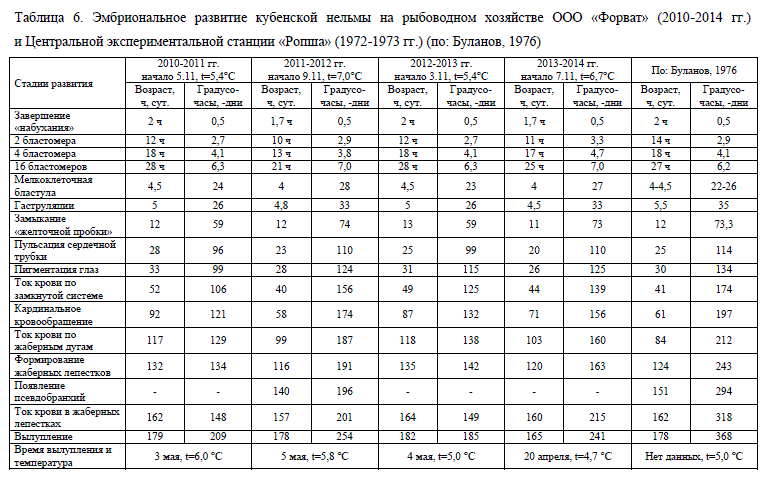
Сравнение эмбрионального развития кубенской нельмы на ООО «Форват» и ЦЭС «Ропша» (Буланов, 1976) также указывает на отмеченные ранее закономерности – высокие инкубационные температуры на ЦЭС «Ропша» ускоряют развитие икры начиная со стадии замыкания желточной пробки (табл. 2). Темп эмбриогенеза в наблюдениях Д.П. Буланова сопоставим с таковым в нашем опыте 2012 г. вплоть до начала пигментации глаз, что объяснимо схожим температурным режимом в этот период развития. Дальнейший температурный режим на ООО «Форват» был существенно холоднее, что сказалось на появлении различий в темпе эмбриогенеза. Начиная со стадии циркулярного кровообращения до начала тока крови по жаберным дугам на ЦЭС «Ропша» потребовалось 43 сут., в то время как в наших наблюдениях в среднем 63 сут. Однако до следующего этапа (формирование жаберных лепестков) в более теплых условиях прошло 40 сут., в то время как в наших наблюдениях в различные годы от 15 до 17 сут.
Подобные особенности развития сложно объяснить, однако в литературе есть сведения, что повышение температуры воды во время эмбриогенеза осенненерестующих рыб, помимо общего ускорения органогенеза способно на определенных этапах тормозить процесс развития (Черняев, 1968; Городилов, 1969). Кроме того, было показано, что высокие температуры на ранних этапах эмбрионального развития байкальского омуля Coregonus autumnalis migratorius, приводят к замедлению его развития на более поздних этапах (Смирнов, 1987). При этом чем выше температура во время прохождения начальных этапов, тем раньше и четче проявляется замедление последующих. Возможно, с этим связано длительное прохождение этапа от кругового кровообращения до кровоснабжения жаберных дуг в наблюдениях Д.П. Буланова (1976), тем более что температурный режим инкубации икры в первые месяцы был достаточно теплым (см. табл. 5).
Помимо перечисленных выше особенностей, отражающихся на эмбриогенезе сиговых, существует ряд других факторов, способных влиять на ход развития зародыша как напрямую, так и косвенно. К первым относятся факторы среды – кроме температуры, это освещенность, pH, гидрохимические показатели.
Ко вторым – генетические особенности рыб, участвующих в нересте, их физиологическое состояние, качество корма в период нагула, качество воды в период инкубации и др. В совокупности перечисленные факторы способны определять достаточную вариабельность эмбриогенеза в пределах одного вида. Тем не менее заключительный этап эмбрионального развития (ток крови в жаберных лепестках) на ЦЭС «Ропша» был отмечен в схожие сроки, что и на ООО «Форват» – 162 сут., но количество тепла, полученное зародышем к этому времени, равнялось 318 градусо-дней, что в среднем на 44% больше, чем на ООО «Форват».
Следовательно, количество градусо-дней, полученное икрой за время инкубации, не коррелирует со скоростью эмбриогенеза. Это дает основание полагать, что на развитие зародышей влияет не сумма накопленного тепла, выражаемая в градусо-днях, а температурный режим на отдельных этапах развития.
В литературе есть немало подтверждений этому факту. На основе экспериментальных работ А.И. Любицкой (1934) и П.Г. Светловым (1934) было установлено, что отношение развивающегося организма к температуре может быть различно на разных стадиях. Снижение скорости эмбриогенеза тресковых рыб (Gadidae) в связи с понижением температуры, приурочено к поздним этапам роста зародыша, в то время как длительность начальных стадий развития изменяется незначительно (Буслов, Сергеева, 2013), что было отмечено и в наших наблюдениях на икре нельмы. Авторы исследования предположили, что подобные особенности развития можно считать адаптацией к вылуплению в наиболее благоприятных условиях.
На неодинаковое отношение развивающегося зародыша к факторам окружающей среды могут также указывать различные регуляторные механизмы этого процесса. Известно, что с развитием эмбриона более примитивные донервные пути регуляции дополняются сначала механизмами нервной регуляции, а к концу эмбрионально-личиночного периода завершается становление нейроэндокринного комплекса (Barton, 2002; Нечаев и др., 2006; Черняев, 2007). Следовательно, в процессе эмбрионального развития наблюдается постепенное подключение к процессам регуляции новых механизмов и изменение роли уже существующих (Лукиянов, 2010).
Таким образом, несмотря на различные инкубационные температуры и количество полученного тепла в процессе развития, эмбрионы рыб одного вида, развивающиеся в различных условиях, к определенному этапу эмбриогенеза могут подходить одинаково развитыми. Как было отмечено ранее, последний этап развития зародыша в икринке (ток крови по жаберным дугам) в разные годы инкубации мы наблюдали в возрасте 157-164 сут., при 149-215 градусо-дней (см. табл. 6). При сравнении наших данных с результатами наблюдений Д.П. Буланова (1976) становится видно, что этот этап развития эмбрионов кубенской нельмы в условиях ЦЭС «Ропша» проходил в том же возрасте – 162 сут, однако количество полученного тепла икринками было существенно выше – 368 градусо-дней.
По-видимому, подобные особенности развития эмбрионов нельмы в данных температурных режимах, как в случае с тресковыми (Буслов, Сергеева, 2013), есть адаптационное приспособление, которое позволяет организму подойти к этапу вылупления наиболее сформированным, но, в то же время, с достаточным эндогенным запасом питательных веществ для дальнейшего развития вне оболочки.
Вылупление предличинок нельмы в 2011-2014 гг., как видно из табл. 6, проходило в первых числах мая в схожем возрасте (178-182 сут.) при прогреве воды до 5,0-6,0°С. Масса предличинок в разные годы равнялась 11,1-11,9 мг.
Выживаемость также находилась в достаточно близких значениях и составляла от 64 до 68%. Однако в этом показателе не была учтена неоплодотворенная икра, которая у сиговых рыб способна развиваться партеногенетически в течение месяца (Борисов, Крыжановский, 1955; Беляева, 1959; Соин, 1968) до стадии замыкания желточной пробки (Черняев, 1982). Количество такой икры нами не определялось.
По срокам вылупления исключением явился 2014 г., в котором выход свободных эмбрионов из икринок произошел 20 апреля при температуре 4,7°С. Продолжительность инкубации составила 165 сут., а масса предличинки была несколько выше, чем в предыдущие годы, и составила 12,4 мг. Несмотря на значительные эндогенные запасы, предлагаемый корм в кишечнике личинок был отмечен в первые дни кормления.
Раннее вылупление в 2014 г. вполне объяснимо динамикой изменчивости температуры воды в период инкубации, которая представлена на рис. 7. Сопоставление графика динамики температуры за период 2013-2014 гг. с данными эмбриогенеза (см. табл. 6) указывает на довольно быстрый прогрев воды, начиная с 130 сут., способствующий ускоренному развитию зародыша. В данном наблюдении был отмечен самый короткий временной отрезок между завершающим этапом эмбриогенеза (160 сут.) и вылуплением (165 сут.). Но, несмотря на это, предличинки были достаточно сформированными, чтобы потреблять внешний корм, что немаловажно при искусственном выращивании рыбы. Более того, раннее вылупление увеличивает вегетационный период, когда идет активное массонакопление быстрорастущей молодью.
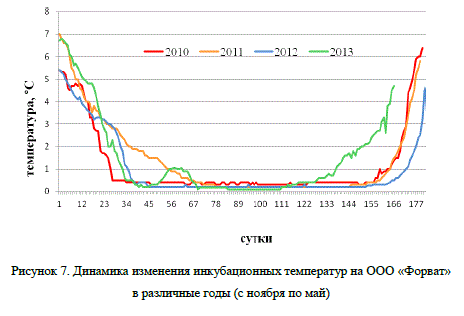
Учитывая указанные выше преимущества, можно заключить, что температурный режим в период инкубации 2013-2014 гг. являлся наиболее предпочтительным для целей аквакультуры, и может быть рекомендован для инкубации икры нельмы в условиях с регулируемым температурным режимом. Для раннего получения предличинок нельмы можно использовать и более высокие температуры. В опыте с икрой сиговых рыб и нельмы (обской популяции) было показано, что значительное повышение температуры (до 8-11°C) на поздних стадиях развития (образование зачатков хвостового плавника и пигментация туловища) приводит к сокращению периода инкубации на 54-55 сут. (Костюничев, 1997). Подобные изменения условий развития существенно не влияют на выживаемость эмбрионов на стадии вылупления, а также на физиологическое состояние предличинок, которые на 3-4-е сут. жизни начинают потреблять внешнюю пищу. Мальковый этап развития такой молоди начинался на 51-52 сут. раньше, чем при естественном температурном режиме.
На широкий диапазон температур (от 0 до 10°C) при котором может проходить нормальное развитие икры сиговых рыб, также указывала О.А. Лебедева (1983, 1985). Однако если повышенные температуры не оказывают отрицательного влияния на поздние стадии эмбрионального развития, то на ранние этапы излишнее тепло действует губительно. У пеляди чувствительный период к высоким температурам определен до стадии пигментации глаз, причем максимум чувствительности приходится на период завершения обрастания желтка икринки (Городилов, 1969). М.Ф. Вернидуб (1950, 1951) были определены критические температуры для сигов на этапе дробления равные 10°C, когда происходит слияние бластомеров. Развитие икры байкальских сигов на ранних этапах при 6°C также значительно увеличивает смертность эмбрионов (Бахарева, 1934). Выживаемость кубенской нельмы, икра которой развивалась при 4,0–8,0°С, составляла всего 1,0–8,0% (Яндовская, Тихонова, 1961). Помимо высокой смертности икры, при вылуплении были отмечены предличинки с различными отклонениями в развитии, которые оказывались нежизнеспособными. Д.П. Буланов (1974, 1979б) также отмечал негативное влияние высоких температур на эмбриональное развитие кубенской нельмы. Выживаемость икры в результате инкубации в относительно низком температурном режиме (tср. = 1,3°С) составляла 68,8% против 57,2% в условиях более высокой температуры (tср. = 2,7°С). Выживаемость икры обской нельмы в диапазоне инкубационных температур 2,5-4,0°C составила 36,9% (Злоказов, 1970).
Учитывая полученные нами результаты и литературные данные по влиянию температуры на эмбриональное развитие нельмы, можно сделать вывод, что начальные этапы эмбриогенеза (включая формирование зародыша и начало органогенеза) должны проходить в условиях «холодного» температурного режима, с понижением температуры от 4-6 до 0,4-0,2°С. В дальнейшем, начиная со стадии окончания пигментации глаз, для получения физиологически полноценных жизнестойких предличинок, возможно повышение температуры вплоть до 8-11°С.
Подобная стратегия зародышевого развития свидетельствует о значительных адаптационных возможностях нельмы, что характерно и для других сигов. На основании работ, проведенных с различными сиговыми рыбами, была выявлена следующая закономерность: эмбриональное развитие осенненерестующих рыб возможно при повышенных температурах в случае, если начальные этапы эмбриогенеза проходят на фоне снижения температуры окружающее среды (Турдаков, Никитин, 1972; Черняев, 2007), Подводя итог проведенным исследованиям по эмбриональному развитию кубенской нельмы, следует отметить существенное влияние внешних факторов среды на этот процесс, среди которых температура занимает ведущее место. Однако другим немаловажным фактором в онтогенезе рыб, в том числе на ранних его этапах, может выступать освещенность.