Формирование полового состава нерестового стада селенгинской популяции байкальского омуля
Вопрос о формировании полового состава
нерестовых стад рыб в литературе освещён
достаточно подробно. Половые же́лезы боль-
шинства пресноводных видов во время заклад-
ки индифферентны: они потенциально двупо-
лы. Определение половой принадлежности
особей возможно лишь при дальнейшем разви-
тии половых желёз. В отличие от высших по-
звоночных, становление пола которых опреде-
ляется генетическими факторами, у рыб этот
процесс связан также с условиями внешней
среды (т. н. фенотипическое или модификаци-
онное становление пола) посредством гормо-
нальной регуляции. На определение пола у рыб
могут влиять такие абиотические факторы сре-
ды, как температура, солёность и освещён-
ность. Происходит это на личиночном этапе
развития [19]. Так, большинство атерин
Menidia menidia, выклёвывающихся ранней
весной при пониженных температурах, стано-
вятся самками, а рыбы, развивающиеся позже
при более высокой температуре, вырастают
самцами [21].
Повреждение эндокринной системы рыб
может стать причиной преимущественного ро-
ждения особей одного пола. Отмечены, напри-
мер, нарушения в эндокринной системе рыб на
ранних стадиях развития под воздействием
вредных выбросов промышленных предпри-
ятий [22] и изменение соотношения полов в
поколениях в сторону преобладания самцов.
Описан случай передифференцировки пола
у баунтовских сигов Coregonus lavaretus baunti
в возрасте 2+. При понижении температуры и
недостатке питания происходит не только за-
держка развития воспроизводительной систе-
мы, но и её перестройка. У самок овоциты за-
мещаются на мужские половые клетки [2].
Дифференцировка пола у молоди байкаль-
ского омуля происходит в возрасте от 3 до 6
месяцев [6]. На процессы гамето- и гонадогене-
за оказывает влияние температура: при её по-
вышении они протекают быстрее.
Для горбуши, размножающейся в реках
Камчатки, свойственно преобладание самцов в
начале нерестового хода. Чем выше числен-
ность нерестового стада, тем сильнее оно вы-
ражено. Если в период массовой миграции на
нерест в реки соотношение полов почти равно,
то в конце нерестового хода на нерестилищах
преобладают самки [10]. В нерестовом стаде
чавычи р. Камчатки отмечено снижение доли са-
мок с 52 % в 1950–1960 гг. до 27 % в 1998 г. [5].
В некоторых популяциях серебряного кара-
ся и видов семейства Poecilidae полностью от-
сутствуют самцы. При отсутствии самцов сво-
его вида развитие икринок активируется спер-
матозоидами других видов рыб. При таком
способе размножения у рыб (гиногенез) потом-
ство состоит из одних самок [20].
Ряд авторов [23; 8] отмечают, что доля са-
мок увеличивается при снижении численности
популяции.
Для принадлежащих к разным морфо-
экологическим группировкам популяций бай-
кальского омуля, который, будучи озёрно-
речным проходным видом, нагуливается в
оз. Байкал, а на нерест заходит во впадающие
реки, характерно определённое соотношение
самцов и самок в нерестовом стаде. Так,
К. И. Мишарин [11] считал естественным такое
соотношение полов в нерестовом стаде наибо-
лее многочисленной популяция омуля, раз-
множающейся в р. Селенге, когда на одну сам-
ку приходится два самца. В нерестовом стаде
омуля из северобайкальских рек Верх. Ангары
и Кичеры в 1962–1968 гг. самцы составляли в
среднем 60–70 % [15]. Доля самцов в нересто-
вом стаде омуля р. Селенги в 1961–1963 гг., по
сравнению с омулем из северобайкальских рек,
изменялась в пределах 45,8–54,0 % [7], а в
1983–1989 гг. в среднем составила 60 % [4].
Лишь в годы, когда на нерест выходит много-
численное поколение омуля, соотношение по-
лов существенно изменяется в пользу самцов.
Популяция байкальского омуля, размно-
жающегося в Селенге, экологически неодно-
родна. Установлен сложный состав и нересто-
вого стада. Выполненное с позиций функцио-
нальной морфологии [1] соотнесение особен-
ностей внешнего строения речных мигрантов с
разнообразием условий их обитания в Байкале
позволило считать, что нерестовое стадо р. Се-
ленги состоит из рыб, принадлежащих к трём
морфо-экологическим группам (морфотипам)
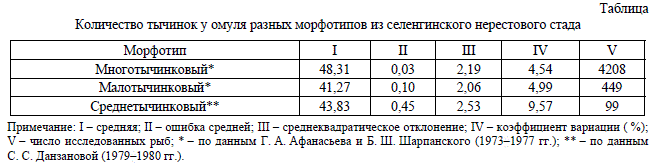
омуля (табл.) [15; 18]: пелагический (многоты-
чинковый), прибрежно-пелагический (средне-
тычинковый) и придонно-глубоководный (ма-
лотычинковый).
Таблица
Количество тычинок у омуля разных морфотипов из селенгинского нерестового стада
Изложенные материалы позволяют сфор-
мулировать цель настоящей работы – выясне-
ние особенностей внутрипопуляционной струк-
туры и формирование полового состава омуля
в нерестовом стаде в Селенге.
Материал и методы
Для решения поставленных вопросов ис-
пользованы данные, характеризующие числен-
ность, размерно-половой и возрастной состав
нерестового стада омуля в 1965–2011 гг., в том
числе собственные материалы 1989–2011 гг.,
собранные в ходе мониторинговых наблюде-
ний за нерестовым стадом омуля в нижнем те-
чении р. Селенги.
В основе определения численности нерес-
товых стад омуля лежит метод перерасчёта на
сутки результатов трёхразовых (утро, день, ве-
чер) ловов жаберными сплавными сетями с оп-
ределением интенсивности суточного хода и
коэффициента уловистости сетей. Разделение
нерестового омуля на морфо-экологические
группы проводили визуально на основе извест-
ных морфологических особенностей последних
[16]. Сбор биологического материала для пела-
гической морфогруппы омуля осуществляли в
соответствии с изменением интенсивности захо-
да, для прибрежной и придонно-глубоководной
морфогрупп набор материала осуществлялся
равномерно в течение всего сезона [4].
Биологический анализ рыб проводился на
свежем материале. Длину тела измеряли с
точностью до 0,1 см от конца рыла до конца
чешуйного покрова и до конца средних лучей
хвостового плавника. Массу тела рыб и массу
гонад определяли с точностью до 1 г. Возраст
определялся по чешуе. Для определения воз-
раста использована методика, учитывающая
особенности формирования годовых зон роста
на чешуе байкальского омуля [17], согласно
которой проведено также переопределение
возраста рыб из проб, хранящихся в фондах
Байкальского филиала ФГУП «Госрыбцентр»,
начиная с 1965 г., что позволило сравнивать
многолетние материалы, собранные разными
исследователями. Всего проанализированы
А. В. БАЗОВ, Н. В. БАЗОВА, В. В. СМИРНОВ
Известия Иркутского государственного универстета
114 266 экз. рыб, из них 23 246 с определением
возраста.
Результаты и обсуждение
В первых числах сентября наиболее много-
численную часть нерестового стада омуля,
входящего в Селенгу, формирует многотычин-
ковый омуль, который принадлежит к нагули-
вающемуся в эпипелагиали Байкала пелагиче-
скому морфотипу.
Во второй половине сентября в реку начи-
нают заходить среднетычинковый омуль при-
брежно-пелагического морфотипа и малоты-
чинковый омуль придонно-глубоководного
морфотипа. По сравнению с первым, они име-
ют меньшую численность. Появление в реке
омуля этих морфотипов наблюдается на фоне
снижения численности многотычинковых рыб
(рис. 1).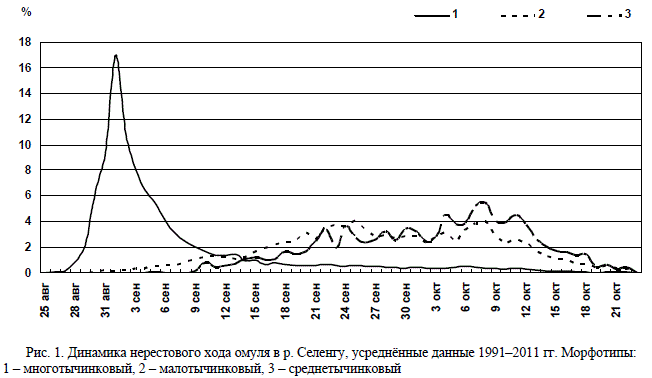
Рис. 1. Динамика нерестового хода омуля в р. Селенгу, усреднённые данные 1991–2011 гг. Морфотипы:
1 – многотычинковый, 2 – малотычинковый, 3 – среднетычинковый
В период наблюдений (1965–2011 гг.) соот-
ношение полов в нерестовом стаде омуля в р.
Селенги изменялось в значительных пределах.
Так, доля самок у омуля многотычинкового
морфотипа составляла в разные годы от 27,94
до 55,70 %, малотычинкового – от 31,58 до
62,50 %, среднетычинкового – от 27,05 до
72,22 % общей численности стада.
Омуль разных морфотипов, заходящий на
нерест в р. Селенгу, различается по срокам по-
лового созревания. Это видно как по возрасту
впервые размножающихся особей, так и по
растянутости сроков созревания рыб одного
поколения. Наиболее раннее созревание (с воз-
раста 5+) отмечено в поколениях омуля сред-
нетычинкового морфотипа. Позже (с возраста
6+) созревает омуль многотычинкового морфо-
типа. Наиболее позднее начало созревания от-
мечено у малотычинкового морфотипа (с воз-
раста 8+). Если в поколениях двух первых мо-
дальными по срокам созревания являются
2–3-возрастные группы, то выход на нерест
поздно созревающего малотычинкового омуля
растянут на 5–6 лет (рис. 2).
Для омуля всех морфотипов характерно бо-
лее раннее половое созревание самцов. В
старших возрастах доминируют самки (рис. 3).
Однако различия между самцами и самками по
скорости созревания неодинаковы в разных
поколениях. Так, если сравнивать отдельные
поколения многотычинкового омуля по сред-
нему возрасту выхода на нерест, в поколениях
1960 и 1981 гг. рождения самцы созревали, со-
ответственно, на 0,9 и 1,0 года раньше самок, а
в поколениях 1987 и 1996 гг. различия между
самцами и самками по скорости созревания
были минимальными (0,1 года).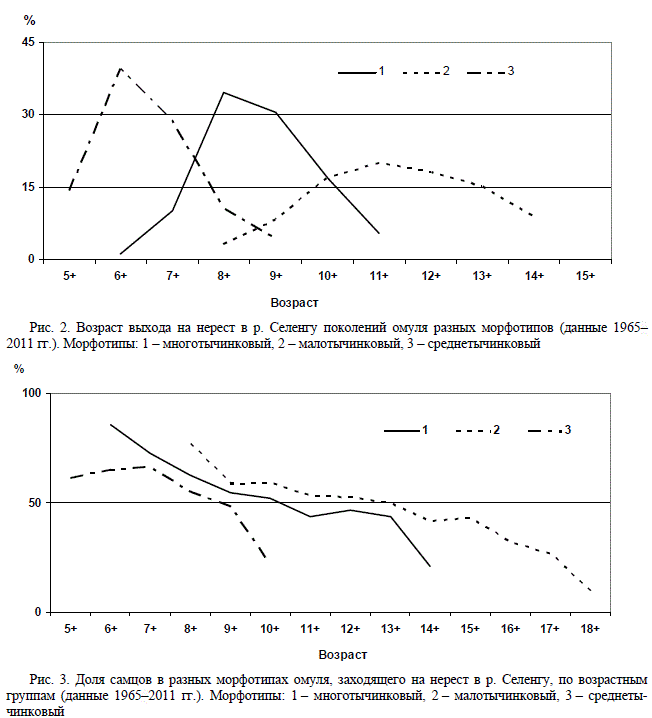
Рис. 2. Возраст выхода на нерест в р. Селенгу поколений омуля разных морфотипов (данные 1965–
2011 гг.). Морфотипы: 1 – многотычинковый, 2 – малотычинковый, 3 – среднетычинковый
Рис. 3. Доля самцов в разных морфотипах омуля, заходящего на нерест в р. Селенгу, по возрастным
группам (данные 1965–2011 гг.). Морфотипы: 1 – многотычинковый, 2 – малотычинковый, 3 – среднеты-
чинковый
Средняя разница в возрасте созревания
самцов и самок многотычинкового омуля за
весь период наблюдений составила 0,38 года. В
отдельных поколениях малотычинкового ому-
ля эти различия достигали 1,6 года, а в среднем
за весь период наблюдений составили 0,6 года.
В поколениях среднетычинкового омуля раз-
личия в созревании особей разного пола изме-
нялись в пределах от 0 до 1,1 года. В среднем
за весь период наблюдений различия между
самками и самцами этого морфотипа составили
0,3 года.
Соотношение полов в нерестовом стаде мо-
гут изменять рыбы, пришедшие на нерест по-
вторно. Выделение таких рыб проводилось на-
ми визуально по состоянию гонад. Как прави-
ло, самцы имеют увеличенные кровеносные
сосуды по сравнению с впервые созревающи-
ми, а у самок в большинстве случаев сохраня-
ются следы предыдущего нереста в виде невы-
метанных икринок, находящихся в полости те-
ла на той или иной стадии резорбции. При
оценке различий в скорости созревания поко-
лений ошибка может быть наибольшей для
среднетычинкового и малотычинкового омуля,
А. В. БАЗОВ, Н. В. БАЗОВА, В. В. СМИРНОВ
Известия Иркутского государственного универстета
у которых доля рыб, пришедших на нерест по-
вторно, составляла в среднем 8 и 10 % соответ-
ственно. У многотычинкового омуля повтор-
ный нерест обнаружен лишь у единичных осо-
бей (0,5 %).
Нами прослежено соотношение полов в по-
колениях многотычинкового омуля 1957–2000 гг.
Во всех поколениях, кроме 1962–1964 гг., вы-
явлено преобладание самцов. Количество са-
мок равнялось количеству самцов или было
несколько больше в 1962 (52,7 %) и 1963 г.
(52,2 %). Соотношение, близкое 1:1, отмечено
также для поколений 1971 и 1982 гг. В то же
время, в некоторых поколениях количество
самцов значительно преобладало над самками.
Так, в генерациях 1972 и 1980 гг. число самок
было минимальным, составив на момент поло-
вого созревания около трети (31,81 и 31,90 %
соответственно) (рис. 4, 5).
Половая структура нерестового стада, точ-
нее, доля самок, принимающих участие в раз-
множении, играет важную роль в формирова-
нии его репродуктивного потенциала. Основа-
ний говорить о том, что какое-либо определён-
ное соотношение самок и самцов является оп-
тимальным для популяции, у нас нет. Вполне
вероятно, что в связи с меняющимися усло-
виями необходимую эффективность размноже-
ния могут обеспечивать различные соотноше-
ния полов в нерестовом стаде. В настоящее
время можно лишь констатировать установ-
ленные за период наблюдений среднемного-
летние соотношения долей самцов и самок: для
многотычинкового омуля таковым является
соотношение 1:1,41 (при колебаниях от 1:0,80
до 1:2,58), для малотычинкового – 1:1,08 (от
1:0,60 до 1:2,17), для среднетычинкового –
1:1,57 (от 1:0,38 до 1:2,80).
В целом на формирование соотношения
полов в нерестовом стаде влияет целый ряд
факторов. Основные из них: избирательное
изъятие промыслом рыб разного пола; возмож-
ность повторного нереста у части особей; раз-
личия в скорости созревания самцов и самок;
различия в среднем возрасте вхождения разных
поколений в нерестовое стадо; изменяющееся
соотношение полов в разных поколениях.
Промысел не играет существенной роли и
может сказаться на соотношении полов только
при нарушении сроков осеннего запрета в при-
устьевых пространствах нерестовых рек. Такая
ситуация наблюдалась в первой половине
1960-х гг. на Селенгинском мелководье в пери-
од перехода на использование промыслом ору-
дий лова с крупной ячеёй. К этому может при-
вести также несоблюдение режима лова в
преднерестовый период в приустьевой части
Селенги и браконьерский лов.
Случаи повторного нереста и связанный с
ними пропуск частью рыб одного–двух нерес-
товых сезонов имеют место во всех популяци-
ях омуля, обитающих в Байкале. Так, нересто-
вое стадо северобайкальской популяции, раз-
множающейся в р. Верх. Ангаре, в среднем на
20 % состоит из рыб, участвующих в нересте
во второй и третий раз [16]. Об этом же свиде-
тельствует существенный процент отнерес-
тившихся рыб в промысловых уловах на мес-
тах нагула в следующем году. По данным
А. А. Войтова [3], на Северобайкальском мел-
ководье особи со следами прошедшего нереста
(синюшные гонады и частично оставшаяся в
полости тела после нереста и резорбирующаяся
икра) составляют до 24,4 %.
В противоположность омулю северобай-
кальской популяции (среднетычинковый мор-
фотип), особи многотычинкового морфотипа
селенгинской популяции со следами прошед-
шего нереста в Байкале на местах нагула
встречаются единично. Это свидетельствует о
том, что в нерестовом стаде селенгинского
омуля многотычинковый морфотип практиче-
ски целиком представлен впервые нерестую-
щимися рыбами. Элиминация отнерестивших-
ся рыб происходит, видимо, осенью, вскоре
после ската из нерестовой реки, так как уже
зимой в контрольных сетных уловах они
встречаются крайне редко. Дальнейшим под-
тверждением этому служит единичное присут-
ствие отнерестившихся многотычинковых осо-
бей во время летней путины на Селенгинском
мелководье. В морфотипах мало- и среднеты-
чинкового омуля р. Селенги «остаток», т. е.
рыбы, выжившие после первого размножения и
пришедшие на нерест повторно, играет более
значительную роль. Почти полная посленере-
стовая гибель омуля многотычинкового мор-
фотипа по сравнению с особями малотычинко-
вого и среднетычинкового морфотипов объяс-
нима экологическими особенностями размно-
жения. Основные нерестилища омуля много-
тычинкового морфотипа, по сравнению со
среднетычинковым и малотычинковым (кото-
рые не поднимаются выше 50 км от устья ре-
ки), располагаются в верхнем течении Селенги
(до 540 км). Не питаясь в реке 1,5–2 месяца и
расходуя на созревание половых продуктов
много энергетических ресурсов, отнерестив-
шиеся многотычинковые омули скатываются
после нереста в нижние участки реки и в зону
влияния речных вод истощёнными и ослаблен-
ными. Таким образом, небольшое влияние на
возрастной состав и соотношение полов в не-
рестовом стаде могут оказывать омули мало- и
среднетычинкового морфотипов, пришедшие
на нерест повторно.
Обилие разнообразных биотопов обитания
омуля в озере создаёт все условия для нерав-
номерного его созревания. Как следствие, на-
блюдаются разнообразие в сроках созревания и
выхода на нерест омуля, принадлежащего к раз-
ным морфотипам, а также неравномерность и
растянутость созревания рыб одного поколения.
Установлены более ранние сроки созрева-
ния самцов по сравнению с самками на протя-
жении всего периода наблюдений. Растяну-
тость полового созревания и неравномерность
созревания самцов и самок, принадлежащих к
разным морфотипам, являются в целом до-
вольно постоянными величинами. Небольшие
колебания в ту или иную сторону могут вызы-
ваться лишь разностью скорости роста в годы с
разными условиями нагула неполовозрелых
рыб в Байкале.
Наблюдаются различия в среднем возрасте
вхождения разных поколений в нерестовое
стадо. Однако трудно утверждать определённо,
вызваны ли они ускоренным созреванием сам-
цов в отдельных поколениях, либо замедлен-
ным созреванием самок.
На возрастной состав и соотношение полов
в нерестовом стаде влияет соотношение полов
в конкретных поколениях, формирующих не-
рестовое стадо. Этот вопрос рассмотрен на
примере омуля многотычинкового морфотипа,
материал по которому был собран в наиболь-
шем объёме. Расчёт показал высокую корреля-
цию (r = 0,79) полового состава рыб в нересто-
вом стаде и слагающих его поколениях (см.
рис. 4).
В связи с вышесказанным, возникает во-
прос о том, как формируется половой состав у
рыб одного поколения. По-видимому, на этот
процесс может влиять характер закладки пола
на первом году жизни и скорость элиминации
рыб в поколениях. Для оценки влияния на де-
терминацию пола абиотических условий были
проанализированы данные по среднегодовому
уровню вод Байкала, численности скатываю-
щихся из нерестовой реки личинок и соотно-
шению половозрелых самок и самцов в кон-
кретных поколениях. Оказалось, что поколе-
ния, появившиеся в благоприятные многовод-
ные годы (1962–1966 и 1983–1988), при выходе
на нерест имели равное количество самок и
самцов, а в поколениях, отродившихся в мало-
водные годы (1972–1982), преобладали самцы
(до 60–65 %) (см. рис. 5).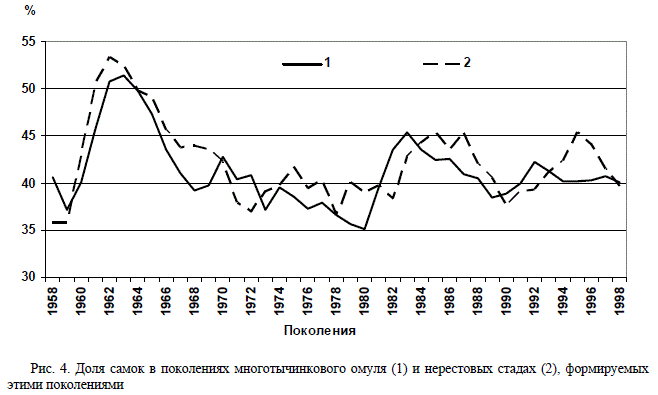 Рис. 4. Доля самок в поколениях многотычинкового омуля (1) и нерестовых стадах (2), формируемых
Рис. 4. Доля самок в поколениях многотычинкового омуля (1) и нерестовых стадах (2), формируемых
этими поколениями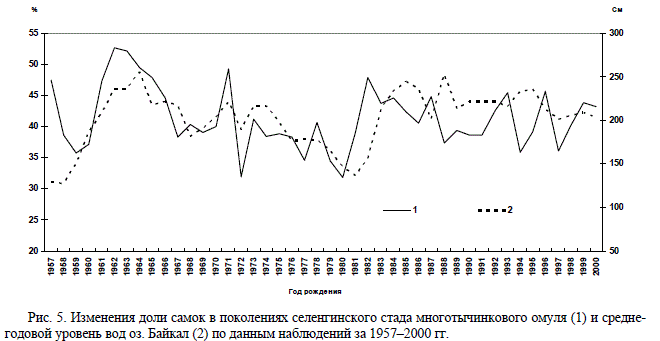 Рис. 5. Изменения доли самок в поколениях селенгинского стада многотычинкового омуля (1) и средне-
Рис. 5. Изменения доли самок в поколениях селенгинского стада многотычинкового омуля (1) и средне-
годовой уровень вод оз. Байкал (2) по данным наблюдений за 1957–2000 гг.
Связи численности скатившихся личинок и
соотношения полов при возврате этого поколе-
ния в нерестовую реку за весь 43-летний пери-
од не выявлено.
Выводы
1. В период 1965–2011 гг. соотношение
самцов и самок в нерестовом стаде омуля
р. Селенги при значительных межгодовых ко-
лебаниях составило в среднем: у омуля много-
тычинкового морфотипа 1,41:1,00; малотычин-
кового морфотипа 1,08:1,00; среднетычинково-
го морфотипа 1,57:1,00.
2. Растянутость сроков созревания много-
тычинкового омуля составляет от 7 до 12 лет,
малотычинкового – от 8 до 15 лет, среднеты-
чинкового – от 6 до 9 лет.
3. У рыб одного возраста в младших воз-
растных группах доминируют самцы, в стар-
ших – самки. Эта зависимость характерна для
всех морфотипов селенгинской популяции
омуля.
4. Доля рыб, пришедших на нерест повтор-
но, составила: у многотычинкового 0,5 %, ма-
лотычинкового 10 %, среднетычинкового 8 %.
5. Доля созревших и вошедших в р. Селен-
гу самок в поколениях многотычинкового ому-
ля в период 1957–2000 гг. колебалась от 31,81
до 52,66 %.
6. Отмечена положительная корреляции
уровня вод Байкала с повышением доли самок
(до 45–55 %) в поколениях омуля, появивших-
ся в многоводные годы.
А. В. Базов1, Н. В. Базова2, В. В. Смирнов 3
1Байкальский филиал ФГУП «Госрыбцентр», Улан-Удэ
2Институт общей и экспериментальной биологии СО РАН, Улан-Удэ
3Байкальский музей ИНЦ СО РАН, Листвянка