Морфометрические признаки пеляди
В ранних публикациях многие авторы увлекались поисками географи ческих закономерностей в клинальной изменчивости признаков, в част ности подчеркивалось закономерное уменьшение числа тычинок и уве личение числа чешуй и позвонков у силовых рыб с запада на восток [Световидов, 1934, 1936; Букирев, 1938; Есипов, 1938; Меньшиков, 1946, 1949, 1951; Кириллов, 1962, 1972; Винглинский, 1966; ?анелі, 1971, 1972; Vladykov, 1934; и др.]. Однако эти выводы были основаны на малом материале, и анализ не охватывал всего ареала вида. При более полном анализе выяснилось, что изменчивость признаков и колебания средних в выборках
связаны не столько с географическим положением Водоема, сколько с температурой, соленостью, химическим составом воды и другими факторами [Решетников, 1980]. Поэтому общие геогра
фические закономерности часто перекрываются под влиянием местных Экологических условий. Поскольку в литературе все еще появляются работы, в которых пытаются выявить «географические закономерности» изменчивости сиговых рыб, остановимся на этом вопросе подробнее на примере пеляди.
При анализе морфометрии пеляди ряд авторов подмечали законо мерности географической изменчивости признаков (Остроумов, 1951; Меньшиков, 1951; Кириллов, 1962; Венглинский, 1966; Канел, 1972; Скрябин, 1979]. Так, на примере пеляди, акклиматизированной в водо емах европейской части СССР, отмечалось уменьшение числа чешуй в боковой линии и числа лучей в непарных плавниках [Андрушайтис, 1963, 1964; ?анеп, 1971, 1972). Чаще всего эти изменения трактовались как проявление общей географической закономерности, причем под черкивалось быстрое изменение признаков по сравнению с родительски ми формами | Крашенинникова, Малышева, 1969; Фролова, 1973; Ка неп, 1972; Скрябин, 1979].
Не отрицая в принципе возможных изменений признаков в новых условиях обитания пеляди, все же отметим, что приведенные выше ра боты были связаны с методической погрешностью: сравнения проводи лось не с родительскими формами из 03. Ендырь-Согомского, откуда ведут свое происхождение все популяции пеляди Европейского Севера и многие южные. В главе 1 уже отмечалось, что до публикации статьи И. С. Мухачева и В. М. Чупретова [1981] в литературе не было данных по морфометрии пеляди из оз. Ендырь, поэтому для сравнения брали данные А. И. Букирева [1938) по пеляди из оз. Нахарвант (бывшее на звание оз. Ендырь-Челикановский вводило в заблуждение). Оз. Ендырь Согомский находится в 30 км западнее г. Ханты-Мансийска, а оз. На харвант расположено в междуречье ?онды и Иртыша (рис. 3). Различия в морфометрических признаках пеляди из этих двух озер весьма су щественны, что и послужило основой для неверных заключений обы стром изменении признаков В НОВЫХ Местах обитания | КрашениННИКО ва, Малышева, 1969; Андрушайтис, 1963, 1964; Фролова, 1973]. Инте ресно, что в водоемах Польши не отмечено отличий в меристических признаках (кроме числа тычинок) у пеляди по сравнению с рыбами из водоемов СССР (Карелия, Литва, Белоруссия, РСФСР) [Mamcari, 1984]. В табл. 3 приведены основные счетные признаки пеляди из этих двух озер и из новых мест обитания.
Подчеркнем еще одну деталь: данные разных авторов по пеляди из оз. Ендырь, полученные через три года (1971 и 1974 гг.), дают досто верные различия по числу чешуй в боковой линии (86,56 и 87,50 при 1=2,93 и р<0,01) и 10 числу лучей в спинном плавнике (9,59 и 9,27 при t=3,40 и р<0,001) (табл. 3). Аналогичная ситуация получается и при
Таблица 3. Меристические признаки пеляди озер Ендырь и Нахарвант

Примечане | Букирев (1938], а Мухачев, Чупретов [1981], а ?анеп (1972) 4 Андрушайтис [1963]; з Фр лова [1973]
сравнении признаков пеляди из Северной Сосьвы (табл. 2): данные в М. Шишмарева (1971 и 1972 гг.) отличаются от данных А. Ф. Пав Това (1975-1979 гг.) по числу тычинок и лучей в Пи А. Таким обра 30м, здесь еще имеет значение и разная система просчета признаков (промеры разными операторами) и изменения средних в популяциях по годам. Поскольку имеются расхождения в подсчете числа чешуй и лучей в плавниках, мы отсылаем читателя к специальной методической работе [Решетников, 1980], а здесь остановимся на основных методи ческих выводах по пеляди.
Меристические признаки. По сравнению с пластическими меристиче ские признаки менее изменчивы. Их окончательное число формируется на первых месяцах жизни (за исключением числа жаберных тычинок) и мало меняется на протяжении жизни. Эти изменения связаны или с селективной смертностью рыб, или со случайностью выборки [Решетни ков, 1980]. Поэтому желательно базироваться на достаточно многочис ленных выборках, собранных за ряд лет. В этом плане наиболее ре презентативными оказались данные, представленные в диссертацион ных работах А. П. Новоселова [1984) и А, Ф. Павлова [1981], на ко торых мы и провели анализ изменчивости признаков упеляди.
Основные меристические признаки пеляди даны в описании вида, который составлен на основании анализа всех доступных данных (табл. 2, 4 и 5). Но все отклонения от описания вида не включены в эти таблицы 1. На рис. 12 приведены пределы колебаний и значения
1 Сомнительными считались данные, которые резко уклонялись от всех остальных Это
в первую очередь относится к числу позвонков илу чей в непарных плавниках, реже кчислу тычинок и чешуй в боковой линии Если будут получены новые достоверные данные, подтверждающие их истинность, то описание должно быть изменено, пока
же мы считаем их сомнительными.
Таблица 4. Меристические признаки пеляди из водоемов европейской части СССР 

Примечание. 1- Суворов [1924], Есипов [1938]; 2 — данные А. А. Козьмина; 3 — Новоселов [1984]; 4 - данные Л. Н. Соловкиной; 5. Сидоров [1974]; f - Пробатов [1938].
Примечание. Использованы следующие оригинальные и опуб- ликованные данные: 1 -- данные А. Ф. Павлова; 2 - Крохалевский [1978]; 3 — Шишмарев (1979, 1985]; 4 -- Бурмакин [1933];
5 — Букирев [1938]: б- Мухачев, Чупретов [19811; 7 --- Коломин [1976); 8 — Красикова [161] и данные
А. А. Вышегородцева; 9 — Бурмакин [1941]; 10 – Попов (1978);
11 — Исаченко [1912], Берг [1948]; 12 — Головков [1973]; 13 - Романов [1985]; 14 -. Логашев [1940]: 15 - Остроумов [1937 ], Букиев
(1938], Решетников (1980]; 16 — Лукьянчиков [1967];
17 — Борисов [1928], Решетников
[198]]; 18 — Венглинский [1966]; 19 -- Кириллов (1972); 20 - ТяITHргянов [1974, 1985]; 21 - Дрягин [1933]; 22 - Новиков [1966); 23 - данные А. А. Ширинского.
средней числа тычинок и чешуй в боковой линии у пеляди из всех во доемов СССР. Прежде всего отметим, что по этим данным невозможно установить географическую закономерность в изменении счетных при знаков. Сравнительно много тычинок встречается у пеЛЯДИ как на за наде ее ареала (Индигские озера, 03. Урдюжское), так и на востоке (озера бассейна Вилюя). Аналогичное явление наблюдается и в числе чешуй в боковой линии (рис. 12).
Анализ 60 выборок пеляди со всего ареала показали, что число ты чинок у нее колеблется от 46 до 69, составляя в среднем 56,54, а число чешуй в боковой линии — от 76 до 102, в среднем 88,62. Для теляди из Водоемов европейской части СССР эти средние значения равны 57,94 тычинки и 86,70 чешуи у речных форм и соответственно 58,00 и 87,70 у озерных. Длят сибирских популяций пеляди эти цифры равны 56,97 и 88,07 для речных и 54,58 и 90,72 для озерных форм. В целом же получается, что у европейских популяций чуть больше тычинок (57,96 по 16 популяциям против 56,05 y 44 сибирских популяций) и чуть меньше чешуй в боковой линии (87,32 против 89,09).
Число чешуй в боковой линии окончательно формируется в первые месяцы жизни и затем у каждой особи остается постоянным (если они не повреждены). Иногда расхождения средних значений по числу че ІШуй обусловлены разными методиками подсчета [ Борисов, 1928; Сыч Аверинцева, 1933]. Обычно принято считать только прободенные чешуи, в противном случае необходимо указать, что приняты в расчет и все прочие. Первой чешуей в боковой линии считается первая краниальная чешуя с канальцем, относящаяся к горизонтальному ряду. Просчиты ваются все чешуи в боковой линии до последней на хвостовом стебле. ?ак правило, у сиговых рыб все чешуи прободенные, но у некоторых особей на хвосте имеются очень мелкие 1—3 чешуйки. Как правило, число чешуй в боковой линии многие авторы счита от правильно, хотя для р. Енисея указано 75 чешуй [Исаченко, 1912], а для 03. Маковск() го — 105 Чешуй [Красикова, 1961 ]. Имеются различия в системе под счета чешуй у американских авторов по другим сиговым рыбам, но по скольку пеляди там нет, то мы их не приводим [Решетников, 1980].
Эксперименты по пересадке сигов из холодных водоемов в теплые иногда сопровождаются снижением числа чешуй [Решетников, 1980; Svardso11, 1970]. У пеляди вновых местах обитания, особенно в водое мах с более теглой водой, может происходить снижение числа чешуй, как это отмечалось в водоемах Архангельской области [Новоселов, 1984), в прудах Белоруссии [Крашенинникова, Малышева, 1969]. Од нако у теляди в псковских озерах [?анеп, 1972] ив озерах Целиноград ской области | Фролова, 1973] произошло увеличение числа чешуй в боковой линии по сравнению с исходой eндырской (см. табл. 3). Анализ возрастной изменчивости этого признака показывает, что прак тически изменений в числе чешуй с возрастом нет (рис. 13), хотя у отдельных поколений могут наблюдаться значительные отклонения от среднего значения (рис. 14, Б, поколение 1971 г.). Эти отклонения (от средних могут быть связаны как с температурным режимом данного года, так и с селективной смертностью рыб данной генерации. Иногда отмечается коррелятивная связь между числом чешуй и средним разме ром рыб в нерестовом стаде сигов [Svardson, 1970].
Число жаберных тычинок с давних пор используется для разделения внутривидовых форм у сигов С. layaretus. Подсчет числа тычинок ве дется на левой жаберної дужке, причем при исследовании рыб малого размера или для многотычинковых сиговых рыб необходимо пользо ваться упой или бинокуляром. Просчитываются все тычинки, в том числе и зачаточные, которые бывают плохо видны, особенно без уве личения. Как и у других видов сиговых рыб с большим число жабер ных тычинок, у пеляди увеличивается число тычинок с ростом рыб.
У пеляди наименьшее число Тычинок встречается на первом году жизни, их число может колебаться от 38 до 50; как правило, редко бы вает более 50 тычинок у рыб моложе двух лет. Поэтому молодь или неполовозрелые особи пеляди желательно не принимать во внимание при расчете средней величины числа тычинок для популяции. На рис. 12 показано увеличение среднего значения числа жаберных тычинок с воз растом у пеляди Печоры и других водоемов Архангельской области. Многие авторы отмечали у пеляди возрастание числа тычинок с возра стом [Букирев, 1938; Есипов, 1938; Меньшиков, 1951; ?анеп, 1971, 1973, 1976; Фролова, 1973; Вершинин, 1984; Новоселов, 1984; и др.]. Проис ходит это за счет увеличения зачаточных тычинок, обычно после второго года жизни число тычинок стабилизируется. Поэтому для вида пеляди в целом мы принимаем колебания числа тычинок от 46 до 69. Указание на число тычинок 40 для бассейна Лены [Кириллов, 1972] относим к разряду сомнительных.
Почти все тычинки у пеляди имеют зубчики [Варпаховский, 1902; Бурмакин, 1953), что позволяет более эффективно использовать их как фильтрационный аппарат. Кроме того, в ротовой полости пеляди на небной части имеется эпидермальная складка, на что впервые обратил


Рис. 12. Вариации числа чешуй в боковой линии (сверху) и числа жаберных тычинок (внизу) у пеляди из водоемов СССР Показаны пределы ?олебаний, 13 - Паднмейские озера;
Внимание Д. Л. Венглинский [ 1966]. Существует определенная связь между числом тычинок и длиной жаберной дуги, более опосредованная связь между размерами тела и числом тычинок в малых озерах Боль шеземельской тундры упеляди намечается явная тенденция к уменьше нию числа тычинок. В новых местах обитания число тычинок может от личаться от числа таковых у родительских форм, что зависит от усло вий питания [Андрушайтис, 1963; Канеп, 1972; Фролова, 1973; Попков, 1979; Новоселов, 1984; Mamcarz, 1971; Peћaz, Hochman, 1984].
, чешуй в боковой линии")
Рис. 13. Изменение числа жаберных тычинок (sp. br.), чешуй в боковой линии (li), позвонков (ut), ветвистых лучей в спинном (D), аналь Ном (А), грудном (P) и брюшном (и) плавниках у пеляди в зависимости от возраста (по данным А. П. Новоселова) 1 – р. Печора; 2 - р Онега, 3 – оз Большое Лебяжье; 4 — Л озеро
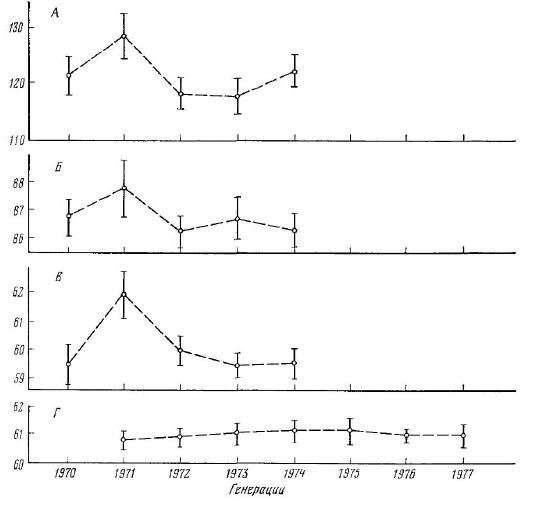
Рис. 14. Изменение числа пилорических придатков (А), чешуй в боковой линии (Б), жаберных тычинок (В) и позвонков (Г) у разных генераций пеляди p. Оби (по данным А. Ф. Павлова) (даны значения средней и ее ошиб?а)
С возрастом пелядь способна питаться все более крупными организ мами, одновременно с этим происходит увеличение расстояния между жаберными тычинками. Так, по данным С. В. Канепа | 1972], среднее расстояние между тычинками (в мм) у пеляди из озер Северо-Запада СССР (185 экз.) меняется с возрастом следующим образом.
Как и другие меристические признаки, среднее число тычинок у пе ляди может меняться в разные годы сбора (табл. 6), что связано со
ые голы сора сталі 11 случайностью выборок (особенно при малом числе рыб) или с влиянием какой-то генерации, которая резко отличается по этому признаку от смежных поколений (рис. 14). Это всегда следует принимать во внима ние при разовых сборах материала.
Число лучей в плавниках у всех сиговых рыб относится к разряду малоизменчивых признаков [Решетников, 1980); скорее всего, значи
Таблица 6. Меристические признаки пеляди Северной Сосьвы и дельты Оби по сборам разных лет (по материалам А. Ф. Павлова; указаны средняя и ошибка средней)

тельные отклонения в числе лучей в парных и непарных плавниках в сборах разных авторов связаны с методической погрешностью. Извест но, что у сиговых рыб окончательное число лучей в плавниках заклады вается к концу первого месяца жизни. В табл. 7 приводятся данные о сроках формирования непарных плавников и жаберных тычинок у пе ляди по материалам Л. В. ?угаевской и Л. Л, Сергиенко [1988). Отме Тим лишь, что эти данные относятся к заводскому выращиванию личи нок, в природе же их развитие протекает быстрее [Богданов, 1982, 1983).
Обычно последние лучи в спинном и анальном плавниках разветвле ны, они считаются одним лучом. Особенно внимательно следует считать неветвистые лучи, поскольку первые жесткие лучи в этих плавниках ма ленькие, плохо видны или скрыты кожей. Подсчет лучей в плавниках лучше проводить с помощью препаровальной иглы. В спинном плавнике пеляди обычно 3—5 неветвистых лучей и 8-12 ветвистых; указание на 2 неветвистых луча или более 5 следует отнести к разряду сомнитель ных данных, которые можно встретить в литературе по сибирской пеля ди. Обычная формула анального плавника III — у 12-----16, указание на меньшее число ветвистых и неветвистых лучей, скорее всего, следует отнести к маловероятным.
Брюшные плавники у сиговых рыб имеют обычно формулу п10, характерную для многих видов сиговых, хотя у пеляди число ветвистых лучей, по данным многих авторов, может колебаться от 10 до 12. От метим, что указание на один неветвистый луч связано с недоучетом,
Таблица 7. Формирование числа лучей в плавниках и жаберных тычинок у пеляди На разных этапах личиночного развития
 поскольку первый луч примыкает ко второму и лежит в несколько иной плоскости Лишь при их разделении иглой или скальпелем удается убе диться, что неветвистых лучей действительно два.
поскольку первый луч примыкает ко второму и лежит в несколько иной плоскости Лишь при их разделении иглой или скальпелем удается убе диться, что неветвистых лучей действительно два.
В отличие от брюшного в грудном плавнике всегда только один не ветвистый луч и около 15 ветвистых (колебания от 13 до 16). Послед Ний самый маленький луч обычно бывает разветвлен, но он считается за один,
?олебания в числе позвонков у пеляди из разных мест обитания представлены в табл. 8, где наряду с данными из опубликованных ра бот представлены оригинальные материалы и наши просчеты числа позвонков у музейных экземпляров (ЗИН АН СССР). Методика подсче та числа позвонков подробно изложена выше (рис. 10). Обычно у пеля ди наблюдаются колебания от 57 до 63 позвонков (редко 64) при зна чении средней для большинства исследованных популяций 60—61.
"Таким образом, все рассмотренные нами меристические признаки пе ляди обладают определенной степенью изменчивости, в порядке воз растания коэффициента вариации (СИ) все исследованные признаки располагаются в такой последовательности: число позвонков= ветви стые лучи в V=Число чешуй в ll-жаберные Тычинки=Число ветвистых лучей в P, A и D=число неветвистых лучей в A и D=число пилориче ских придатков (табл. 9). Как и у сиговых рыб, степень изменчивости признака зависит от сроков его закладки [Решетников, 1980]. Не слу чайно, что к наименее изменчивым признакам относится число позво ков, так как их окончательное количество формируется еще в эмбрио генезе, а наиболее изменчивым оказалось число пилорических придат ков, которое постоянно возрастает во время жизни рыбы и связано с особенностями питания рыб. Обычно коэффициенты вариации большин ства признаков не превышают 10%, что свидетельствует об однородно сти выборок и небольших погрешностях в промерах. Однако при малых значениях средних величин (неветвистые лучи в риА) коэффициенты вариации могут быть более 10% [Решетников и др., 1975; Решетников, 1980).
с нашей точки зрения, проведенный анализ меристических призна ков позволяет сделать следующее заключение. Эти признаки можно ис пользовать в таксономических и популяционных исследованиях, если сборы охватывают рыб разного возраста и выборки достаточно велики или если пробы собирались в течение ряда лет. В этом случае возмож ные небольшие различия средних значений у рыб разных поколений (генераций) нивелируются. Полезным может оказаться приВтечение методов из теории информации и использование ЭВМ
Пластические признаки Многие авторы отмечали, что в каждом во доеме пелядь обладает своей характерной формой тела, несколько от личной от других. Частично это объясняется тем, что в каждом водоеме живут рыбы с разным темпом роста.
У пеляди в возрасте 1 - 2 мес еще четко выражены ювенильные чер Ты, для рыб характерны все черты инфантильного организма: относи тельно велик размер готовы, глаз, верхней челюсти, хвостового стебля
Таблица 8. Число rіозвонков у пеляди в водоемах СССР
** Данные автора уменьшены на один позвонок согласно Ю. С. Решетникову [1980].
* Просчеты Ю. С. Решетникова по коллекциям ЗИН АН СССР;
Таблица 9. Коэффициенты вариации (со) меристических признаков у пеляди из разных водоемов СССР
 меристических признаков у пеляди из разных водоемов СССР")
и хвостового плавника. Для мальков обычны невысокий спинной плав ник, небольшой анальный плавник и типично элипсовидная форма тела. Утрехмесячной пеляди относительно уменьшается дистальная часть тела за спинным плавником, особенно хвостовой стебель. В возрасте 6-— 12 мес пелядь приобретает черты взрослой рыбы и по внешнему виду мало от них отличается. Однако аллометрический рост многих призна ков продолжается и в дальнейшем. Основной тип возрастных изменений в пропорциях тела связан с уменьшением размеров головы и глаз, суве личением высоты тела и большей вальковатостью брюшка [Березов ский, 1924; Борисов, 1928; Есипов, 1938; Меньшиков, 1949, 1951; ?анеп, 1971, 1976; Шишмарев, Лугаськов, 1977; Решетников, 1980; Павлов, 1981; Завьялова, 1984; и др.].
Это связано с тем, что по мере роста рыбы не все части тела растут пропорционально изменениям длины (изометрия), поэтому наблюдается положительная или отрицательная аллометрия пластических признаков. I lочти всегда относительные размеры головы и глаз характеризуются отрицательной аллометрией, а наибольшая Высота тела — Положитель ной аллометрией (рис. 15). Отметим, что это относится не ко всем по пуляциям пеляди и наличие изометрического или аллометрического ро ста пластических признаков специфично для каждой популяции. Так, антедорсальное расстояние у большинства сиговых рыб характеризует ся изометрией, в том числе и для пеляди из водоемов Ленинградской области [?анеп, 1976], однако у разных популяций пеляди из водоемов Архангельской области этот признак обладает положительной алломет рией (рис. 15 ІІІ). И такие признаки, как длина хвостового стебля, раз меры парных и непарных плавников, имеют самые разные типы алло метрии. Характер изменения признаков специфичен для каждой попу
Морфометрические признаки
, антедорсального расстояния (ІІ), наибольшей высоты тела")
Рис. 15. Изменение длины толовы (I), антедорсального расстояния (ІІ), наибольшей высоты тела (III), высоты спинного плавника (IV) и длины брюшного плавника (V) у разных популяций пе ляди в зависимости от раз меров тела (по данным А. П. Новоселова) 1 — пелядь р Печоры; 2 — пелядь р Онеги; 3 — лелядь 03 Б Лебяжье; 4 - пелядь Андозера
Поясним это на примере пеляди из водоемов Архангельской об ласти: при изометрическом изменении этого показателя у трех популя ций пеля Ди у печорской пеляди имеется явная положительная алломет рия (рис. 15.V). В то же время при слабой отрицательной аллометрии высоты спинного плавника у всех четырех популяций пелядь р. Печоры отличается по этому признаку от всех остальных популяций при сравне
нии всех размерных групп (рис. 15.1V).
?ак уже отмечалось, тугорослые формы пеляди сохраняют черты мо ЛОДЫХ особей и IOэтому обладают большими относительными размера ми головы и глаз. Иногда разная направленность возрастных изменений признаков позволяет установить принадлежность рыб к разным локаль ным стадам [Смирнов, 1969; Смирнов, Шумилов, 1974]. Но бывают и противоположные заключения. Так, анализ возрастных изменений при знаков у мелкого сига-нельмушки из оз. ?убенского и крупного сига казарки позволил установить, что крупные сиги представляют собой не (отдельную форму или стадо, а являются просто старшими по возрасту особями сига-нельмушки [Лебедев, 1975].
Довольно часто в литературе можно встретить попытку объяснить те или иные различия в пластических признаках их приспособительным Значением в определенных условиях обитания [?анеп, 1976; Крохалев ский, 1978, а, б; и др.]. Однако все признаки взаимосвязаны, образуя как бы плеяды признаков [Андреев, Решетников, 1977]. Эти корреля тивные связи приводят к тому, что изменение одного признака неиз бежно приводит к изменению других. В настоящее время уже никто не принимает, что каждый признак контролируется определенным геном, поэтому из менения в структуре гена могут привести к изменению целого ряда признаков (сцепленные признаки). В этом плане наши стремления объяснить изменения признаков приспособлением к новым условиям обитания выглядят наИВНЫМИ и субъективными.
Если брать пелядь в разные сезоны года, то можно выявить изме нения в признаках, связанные с изменением внешнего облика созреваю щих самок и самцов [Крохалевский, 1978, б]. Ярко выраженного поло вого диморфизма у большинства видов сиговых рыб нет (Решетников, 1980), за исключением тех случаев, когда пробы берутся перед самым нерестом. Такие различия между пола ми перед нерестом отмечены и у пеляди. Обычно они выражаются в изменении антедорсального пост дорсального, пектровентрального и вентроанального расстояний [Беля ев, Венглинский, 1976; Крохалевский, 1978а; Попков, 1979; Вершинин, 1984; и др.].
Наиболее существенно меняются пластические признаки пеляди при ее вселении в новые водоемы. Есть основания полагать, что у всех сиго вых рыб внешний вид рыбы (экстерьер в большей степени контроли руется условиями жизни, чем передается по наследству от родителей
Решетников, 1980). Можно привести довольно много примеров, когда генетически однородные особи при выращивании в новых местах обита ния имели разный темп роста и в конечном итоге превращались в рыб, совершенно непохожих друг на друга. Имеется много подобных выводов и по пеляди при анализе ее признаков в новых местах обитация [Андрушайтис, 1963, 1964; Волкова, 1962; Крашенинникова, Малыше ва, 1969; Волошенко, 1973; Фролова, 1973; Горбунова и др., 1976; Ники тин, 1977; Полков, 1976; Вершинин и др., 1979, 1981; ?арасев, 1980;
Жданкина, Поляков, 1982; Завьялова, 1984; Новоселов, 1984; Верци НИН, 1984; Романов, 1985; Новоселов, Решетников, 1988; и др.]. Если в новых местах обитания у пеляди меристические признаки меняются слабо, то ІПластические признаки чаще всего меняются, и притом значи тельно. Поскольку большинство популяций пеляди в новых местах оби тания ведет свое происхождение от пеляди из 03. Ендырь, приводим данные по морфометрии пеляди из этого водоема [Мухачев, Чупретов, 1981] (табл. 10).
Таким образом, использование пластических признаков в качестве видового или внутривидового критерия для выделения отдельных попу ляций или стад осложняется многими методическими трудностями (учет размерной и возрастной изменчивости, изменение в зависимости от темпа роста). Хотя в некоторых случаях использование морфометрии в популяционных исследованиях давало положительные результаты, Когда разные стада ому 21я обладали разными типами аллометрического роста [Смирнов, 1969; Смирнов, Шумилов, 1974] или когда с помощью этого метода удалось установить единство крупных и мелких особей сига в ?убенском озере [Лебедев, 1975]. Новые возможности открыва ет использование современных методов математического анализа с применением ЭВМ | Андреев, Решетников, 1977, 1978; Решетников, 1977б, 1980, 1981; Андреев, 1980, 1987]. Однако использование новых методов и приемов не разрешает всех трудностей проблемы, но эти ме Тоды оказываются мощным и необходимым средством обобщения иil формации. Для биолога же главным остается снабжение машины доб ротной исходной информацией, чтобы материал был собран без методи ческих просчетов и грубых методических ошибок.
Интерьерные признаки, или морфофизиологические показатели, в физиологических и популяционных исследованиях используются та кие IIоказатели, как относительный вес глаз, мозга, сердца, почек и пе чени. Применяются они и при исследовании популяционной структуры пеляди [Смирнов и др., 1972; ?анеп, 1976; Шишмарев, 1976; и др.]. Однако этот метод не нашел широкого применения в ихтиологии. Более "Гого, имеются серьезные критические за мечания в его адрес, поскольку его применение началось без должной методической разработки. Мето дика морфофизиологических показателей не относится к разряду обще принятых ихтиологических методик, поэтому результаты работ, выпол ненных с ее применением, мы не рассматриваем. Замечания, высказан Hые в адрес пластических признаков, в равной мере применимы и к морфофизиологическим показателям.
Из всех морфофизиологических показателей следует выделить вес печени. Этот индекс малоприменим в исследованиях по ВІТутривидовой структуре вида сигов или при популяционных исследованиях, посколь ку его сезонные изменения, связанные с созреванием гонад и нагулом
Таблица 10. Морфометрические признаки пеляди оз. Ендырь-Согомский [по: Мухачев, Чупретов, 1981]


4, 19 н— наибольшая высота тела, 1А — длина сенезвания анального h-— наименьшая высота тела,
Главинка, ар - антедорсальное расстояние, ?А — высота анального плавника, pp - Гостдорсальное расстояние, IP --- длина грудирrо плавника, аў - антевентральное расстояние. Ту — длина брюшного лларинка,
- антеаїнальное расстояние, р-у — тектовентральное расстоя р! - длина хвостового стебля,
-- длина основания слинного -A --- Вентрзанальное расстояние,
Плавинка,
po — ?атла?ничное расстояние, - высота спинного плавника, ізіх — высота верхнечелюстной
Условные обозначения,
r — длина рыла, o -- горизонтальный диаметр
глаза, hС — высота головы у затылка,
с - длина головы, io -- ширина лба, lerix — длина верхнечелюстной кос
Ние,
Ти,
{md - длина нижней челіости,
рыб, намного перекрывают различия между популяциями Решетников и др., 1970; Решетников и др., 1971; Решетников, 1980]. Индекс печени служит надежным критерием для выделения созревающих самок, а также для выделения рыб, пропускающих нерестовый сезон.
Ю. С. Решетников, А. К. Козьмин, А. П. Новоселов, А. Ф. Павлов