Состав ихтиофауны северных и высокогорных озер
Состав ихтиофауны европейских северных водоемов и субальпийских озер однообразен: она представлена 10— 15 видами рыб, из которых преобладают сиговые, лососевые, довольно часто встре
1 Ценность этого показателя широко обсуждалась на двух междуна родных симпозиумах: «Лососевые сообщества в олиготрофных озерах - Salmonid communities in oligotrophic lakes (SCOL)».- J. Fis. Res. Bd. Cana da, 1972, v. 29, N 6и «Окуневый международный симпозиум - Percid In ternational Symposium (PERCIS)».-- J. Fish. Rеѕ. Ed. Canada, 1977, v. 34, N 10.
чается хариусовые, колюшки, щука, налим; небольшим числом видов представлены окуневые и карповые. Более сложный состав ихтиоценозов — в водоемах Сибири. В арктических и субарктических. пресноводных водоемах сиговые рыбы составляют основную долю ихтиопродукции, давая 70—80% общей добычи рыб. Так, в арктических озерах Канады и Фенноскандии основу уловов дают озерный голец и сиг (до 90%), причем численность и биомасса каждого из этих двух видов находятся в обратной зависимости. В последние годы популяции сига в шведских озерах количественно доминируют над популяцией гольца [Svardson, 1976].
Здесь уместно отметить общие тенденции развития экосистем . Экосистема — это взаимосвязанное сообщество организмов, кото рые взаимодействуют с физической средой таким образом, что соз дают четко выраженный поток энергии, трофическую структуру и круговорот веществ внутри системы [Одум, 1975]. Это незамкну тая система со стабильной структурой во времени и пространст ве, это те «блоки», из которых состоит вся биосфера. Любая эко система имеет свою историю развития и эволюцию. Развитие эко системы, часто называемое экологической сукцессией, идет «по правилу максимума энергии в биологических системах», т. е. со сдвигом потока энергии в сторону увеличения количества энергии, направленного на поддержание системы.
В зрелых экосистемах наблюдается большое видовое разнооб разие, хорошо выражена специализация по пищевым нишам, име ются длинные и разветвленные пищевые цепи и длинные жиз ненные циклы. По сравнению с молодыми, развивающимися си стемами в зрелых экосистемах отношение продукции к биомассе (Рів) невелико, а отношение продукции к обмену приближается к единице (PIR=1).
Теоретически в зрелых системах высокая биомасса поддерживается поступающим потоком энергии, достигая максимума на зрелых стадиях, и, как следствие этого, чистая продукция сообщества (в нашем случае и рыбопродукция) вели ка на ранних стадиях развития экосистемы и мала или практически равна нулю в зрелой экосистеме. По многим показателям девственные северные озера приближаются к зрелым экосистемам. Можно сказать, что лососево-сиговые олиготрофные озера — это климаксное, зрелое состояние водных экосистем Севера. Это со стояние преобладает и будет преобладать при естественном состо янии водного режима, при данном уровне поступления биогенов и данном энергетическом потоке. Обычно все изменения в приро де имеют тенденцию к образованию устойчивых экосистем при высокой биомассе и малой продукции. Эта закономерность часто находится в противоречии со стремлением человека иметь высокий ,
1 Понятие «экосистема» очень близко понятию «биогеоценоз» [Сукачев, 1967], однако мы пользуемся понятием «экосистема», которое более приня то в ихтиологии и экологии. Для характеристики состава рыбного сообще ства и системы отношений среди рыб применяется термин ихтиоценоз.
выход продукции при небольшой ее биомассе, что характерно для молодых, незрелых систем типа пашни или луra Дажо, 1975; Одум, 1975].
Общепризнано, что устойчивость и сложность экосистем за висят от разнообразия составляющих ее видов и функциональных связей. Сложность северных экосистем достигается не столько увеличением числа видов (их там немного), сколько обилием раз личных экологических форм, которые в биоэнергетическом отношении эквивалентны разным видам. Поэтому большое разнообразие форм у сиговых и лососевых рыб можно рассматривать как резуль тат развития северных экосистем по пути усложнения связей.
Обычно первичная продукция арктических и субарктических озер невелика и составляет около 8 г С/м? в год. Так, первичная продукция озер ?ольского полуострова и озер Большеземельской тундры за вегетационный период составляет не более 200 ккал/м* при месячной продукции фитопланктона 7,7 гC/м”, или 60— 77 ккал/м”, и его биомассе 1,2—1,9 ккал/м”, или 0,19 гC/м2 [Большие озера Кольского полуострова, 1975; Винберг, 1976]. Однако при низких значениях первичной продукции имеющаяся в озерах ихтиомасса может быть достаточно высокой (до 50 кг/га и более), что свидетельствует о низкой скорости потока энергии. Значения P/B-коэффициента фитопланктона для 4 северных озер составляет 86— 110, для бактериопланктона — 30 — 40, зоопланкто на — 2— 13 и бентоса — 1,1 — 3,0 [Винберг, 1976]. К сожалению, в ихтиологии очень мало данных о продукции рыб, поэтому приходится оперировать уловами и рассчитывать условный PB-коэффициент по количеству выловленной рыбы. Обычно принимается, что вылов составляет значительную или даже большую часть общей годовой продукции ихтиомассы промысловых озер. Для ряда водоемов вылов оценивается долей порядка 0,30—0,70% от первичной продукции [Винберг, 1976]. По нашим расчетам на основании материалов Т. Т. Болотовой [1977a], Р'/B-коэффициент для сига-лудоги равен 13%. Для севанских сигов P/B-коэффициент равен 25%. Оценка ихтиомассы 95 озер Балтийского бассейна севернее 60° с. ш. после их обработки ихтиоцидами колебалась от 3 до 200 кг/га, составляя в среднем 31:3 кг/ra [Китаев, 1977); близкие числа приводятся и для озер Канады: 11—80, в среднем 32 кг/га, т. е. это оценка всей имеющейся ихтиомассы в данный момент.
Ежегодные стабильные уловы для большинства водоемов Северной Карелии до бурного развития любительского рыболов ства (до 1955 г.) составляли 2—5 кг/га, и грубый подсчет P/B-ко эффициента дает 16—30%. Это подтверждает тезис о том, что значительная часть энергии идет на поддержание имеющейся биомассы.
Сукцессии в олиготрофных экосистемах происходят под влия нием климатических факторов (стойкие и длительные изменения температуры и водности) или под влиянием человеческой деятель
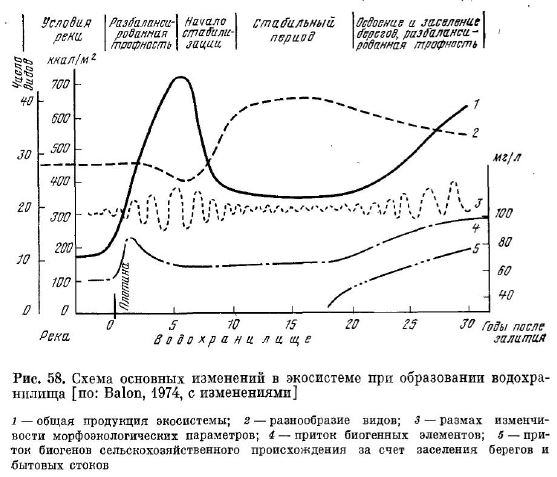
Рис. 58. Схема основных изменений в экосистеме при образовании водохра НІІлища [по: Balon, 1974, с изменениями
1 — общая продукция экосистемы; 2 — разнообразие видов; 3 — размах изменчивости морфоэкологических параметров; 4 — приток биогенных элементов; 5 — при ток биогенов сельскохозяйственного происхождения за счет заселения берегов и бытовых стоков
ности: 1) создание водохранилищ; 2) неправильное ведение рыбо ловства в течение ряда лет, которое расценивается как стрессовый фактор; 3) вселение новых видов рыб; 4) изменение условий оби тания путем загрязнения водоема, вырубки лесов на площади водосбора и т. п. Пример такой сукцессии можно видеть при соз дании водохранилища, которая в схематическом виде изображена на рис. 58. Сначала происходит резкое увеличение изменчивости, темпа роста и биомассы всех живых компонентов системы (в том числе и рыб). Затем наступает снижение изменчивости, падение биомассы и переход системы в стационарное состояние на новом уровне. В отношении сиговых рыб хороших примеров нет, поэтому остановимся подробнее на анализе трех других факторов (про мысел, акклиматизация, эвтрофирование).