Изменчивость меристических признаков у сиговых рыб
По сравнению с пластическими счетные признаки менее изменчивы, однако в оценке их значимости в экологических и таксономических исследованиях существуют самые разные точки зрения. Многие авторы подчеркивали географическую закономерность изменений меристических признаков, в частности уменьшения числа тычинок и увеличения числа чешуй и позвонков у
 ІІ длины нижней челюсти")
Рис. 2. Средние показатели наименьшей высоты тела (1) ии длины нижней челюсти (2) (в % от 7Лины тела) у полупроходных (А), речных нозерно речных (Б) п (зерных (В) сигов
Цифрами на оси абсцисс обозначены следующие водоемы: 1 — р. Воронья; 2 -- р. Ту лома; 3 - - p. Поной; 4 — р. liемь; 5 — реки Белого моря; 5 — р. tiара; 7 -- ). Юрия- . бей; 8 — р. Сибирча; 9 — р. Обь, низовья; 10 — р. Обь; 11 — р. Тыда; 12 --- p. E11 II сей, устье; 13 — р. Енисей; 11 — р. Оленейи; 15 — р. Лена; 16 — р. Хрома; 17 — р. Ко лыма; 18 -- р. Анадырь, с{?-востряк; 19 — р. Нярка (ко:Tьский полуостров); 20 - 03, Подпахтинское; 21 — р. Вонка (Карея); 22 — р. Северная Двина; 2.3 — реки Те чора Усть-111.Тьма; 24 -- реки Печора, Уса; 25 — 1). Уса; 26 -- р. Err исей; 27 — оз, УГеликое (бас. р. Пясина); 28 — р. Ангара; 29 — Братское водохранилище; 30 — рекин Северного Байкала; 31 — р. Баргузин; 32 — р. Вилюй; 3.3 — р. Лена; 341 — р. Чара, бассейн Лены; 35 — р. Аналиырь (сиг-горбун); 36 — 2. Пенкина; 37 — оз. 11андра, Нестеров о-в; 38 — 03. IIмаtидра, губы ППеча | Белая; 39 - 03. Осинов ское; 10 — Iунозеро; 41 — Охтозеро; 49 — Чингльс-вр; 4.3 — Кензис-вр (мало тычинковый сиг); 44 — Кензис-авр многотычинковый сиг); 45 — Пяозеро; 46 — Топ озеро; 47 — Вингьозеро; 48 — иКеретьозеро; 19 — 03. Кета; 10 — 03. Мелкое; 51 -- 03. Таймыр (высокотелый сиг); 52 — 03. Таймыр (низкотелый); 13 — 03. Телецкое; 54 — озера Верхнего Енисея; 55 — 03. Леприндо, бассейн Лены
сиговых рыб с запада на восток (Световнидов, 1934, 1936; Буки рев, 1938; Есипов, 1938; Меньшиков, 1946, 1949, 1951; Суворов, 1948; Кириллов, 1967, 1972; Vladykov, 19:34]. Впоследствии было установлено, что это связано не столько с местоположением Водоема, сколько с температурой, соленостью и некоторыми другия ми факторами, поэтому общие географические закономерности часто перекрываются влиянием местных экологических условий. Многочисленными экспериментами и наблюдениями в природе на примере Могих видов рыб подтверждена зависимость меристических признаков (от "усиtувий впешg:й студыи, її в первую оче редь от температуры, света, солености [Татарко, 1968; Gabriel, 1944; Taning, 1952: Lindsey, 1962а; ииemper, Blaxter, 1961]..
Поскольку в литературе встречаются расхождения в методике подсчета числа тучей в плавниках, позвонков и чешуй в боковой иННи Ни, то следует описать нашу систему, которой мы пользо вались и к которой старались привести опубликованные данные других авторов.
Число лучей в плавниках. Число лучей в парных и непарных плавниках у сиговых рыб мало изменчиво, часто одно и то же число лучей характерно дия разных видов рыб, а небольшие различия средних значений перекрываются методическими погрешностями счета разных авторов. У сиговых рыб лучи в плавниках начинают закладываться на ранних этапах личиночного развития и обычно к концу первого месяца жизни формируется око
чательное число лучейи [Европейцева, 1949; Смольянов, 1957], в которое не меняется с возрастом.
Отметим, что обычно последние лучи в спином и анальном плавниках разветвлены, они считаются одним лучом. Особенно внимательно следует считать неветвистые лучи, поскольку тер Вые лучII в ри Аочеть маленькиrе и плохо ВИДиНЫ. Подсчет лу
чей в плавниках лучце проводить с помощью препаровальної тиглы. Брюные плавники у сировых рыбо, как правило, имеют
два неветвистых луча, причем первый луч лежит в другой пло скости, поэтому многие авторы его пропускают при подсчетах. Обычная формула брюшного плавника | 10 | грудного и 15 с небольшими отклонения и повторяется у многих видов сиговых рыб.
Число позвонков. Число позвонков зависит от числа мноме ров, их окончательное число определяется на ранних эмбрио Анатьных этапах развития II не меняется с возрастом, в полевых
условиях чисто позвонков мы просчитывали на светих рыбах: рыбу разрезати нонком вдоль позвоночника ночищали края поз - вонков от мышц. При известном навыке ошибка практически
исключена можно получать массовый материал. У музейных 1 экземпляров позвонки просчитывали по рентгеноснимкам. По скольку нет единообразия в системе подсчета позвонков у сиро Вых рыб, то на методике подсчета стоит остановиться подробнее.
У сиговых рыб первый позвонок срастается с черепом и его
19
трудно отчленить; обычно этот рудиментарный позвонок не учи тывается [Покровский, 1967; Шапошникова, 1968; Решетников и др., 1975]. Первым считался такой позвонок, который имел два сочленения (справа и слева); этот четкий признак всегда позволя ет фиксировать начало отсчета. Последние три позвонка у всех сиговых рыб загнуты вверх, причем самый последний позвонок имеет только одно, проксимальное сочленение и в отличие от всех остальных характерную форму флажка или вымпела (рис. 3). Из последнего позвонка выходит мягкий уростиль, который не учитывается при подсчете числа позвонков.
обычно колебания числа позвонков у определенного вида не превышают 5—6. Приводимые иногда в литературе пределы ко лебании числа позвонков порядка 10-15 для нельмы, чира, сига, тугуна, пеляди и муксуна из водоемов Сибири [Новиков, 1966; Карантонис и др., 1956; Кириллов, 1972; Пилин, 1974] следует считать сомнительными и нуждающимися в проверке, так как зни противоречат всем другим известным данным. Даже у мурен при колебании числа позвонков в семействе от 120 до 180 пределы колебаний у одного вида не превышают 8-9 позвонков | Mc-Cleneghan, 1976].
Наши подсчеты числа позвонков усига Чунозера не выявили Никакой коррелятивной связи между средним числом позвонков в поколении и температурой воды во время инкубации икры в естественных условиях (с октября по апрель). Не отмечено так же закономерных изменении числа позвонков у сигов разных по колений (рис. 4) и у рыб разного возраста (табл. 1). Незначи тельные колебания средних вызваны малыми выборками или случайными отклонениями средних. Поэтому мы полагаем, что подмеченные В. Я. Ермохиным [ 1974] изменения числа позвонков имеют характер фенотипической изменчивости, представляют собой частный случай и не имеют направленного характера, как считал этот автор. Сборы разных лет практически дают идентичные результаты по всем меристическим признакам, если их оценки базируются на достаточно многочисленных выборках (табл. 2).
Число чешуй в боковой линии окончательно формируется в первые месяцы жизни и в течение жизни у каждой особи остает- . ся постоянным (если они не повреждены). Иногда расхождения средних значений этого признака обусловлены разными методиками подсчета (Борисов, 1928; Сыч-Аверинцева, 1933]. Обычно принято считать только прободенные чешуи, в противном случае необходимо указывать, что приняты в расчет и все прочие. Пер вой чешуей в боковой линии считается первая краниальная чешуя с канальцем, относящаяся к горизонтальному ряду. Просчитываются все чешуи в боковой линии до последней на хвостовом стеб ле. Как правило, у сиговых рыб все чешуи в боковой линии прободенные. Но у некоторых особей на хвосте имеются очень мелкие 1 — 3 чешуйки без пор. В наших данных эти мелкие чешуйки в 
Рис. 3. Схема просчета краниальных и каудальных позвонков у сиговых рыб Цифрами обозначены позвонки; с — ребро; н., ..., Н, — гитупалии; u — уростиль; осе. 1. — боковая затылочная кость; D. осе. - основная заты,точная кость; R — руди ментарный позвонок
Рис. 4. Изменение числа чешуй в боковой линии (I), позвонков (II) и жа берных тычинок (1) у разных генераций сигa Чунозера Показаны среднее значение и ошибка средней
расчет не принимались (но отмечались на карточках). Отметим, что некоторые американские авторы считают чешуи только до конца позвоночника (Dymond, 1943 ], другие считают все чешуи, в том числе и мелкие на хвосте [Lindsey, 1962b ]. Отсюда могут быть расхождения средних величин.
Многие авторы отмечали изменения числа чешуй у разных популяций в зависимости от условий обитания. Часто северные популяции сигов имеют больше чешуй в боковой линии и соот ветственно более мелкую чешую, чем южные. Эксперименты по пересадке сигов в Швеции из холодной реки Индaльсельвен в пруды с более теплой водой показали, что в новых условиях потомство имело меньше чешуй в боковой линии, чем их родители: в реке у сигов было 89— 110, в среднем 97,0 чешуй, ав прудах — 80 — 93, в среднем 85,7 чешуй в 1 [Svardson, 1970]. Свердсон | Svardson, 1952] отмечал также тесную коррелятивную связь между средним числом чешуй в у каждой популяции и сред ним размером рыб в нерестовом стаде: для 94 шведских популя ций коэффициент корреляции (r) между этими величинами ра вен 0,7004 при р<0,001.
Наши дан?ые показывают, что практически со второго года жизни число чешуй в боковой линии учунозерских сигов остается постоянным, не зависит от года сбора материала (см. табл. 2) имало меняется от поколения к поколению (см. рис. 4), т. е. при неизменных условиях обитания число чешуй в боковой линии — достаточно стабильный признак для каждой популяции сигов.
Число жаберных тычинок с давних пор используется при раз делении видов и внутривидовых форм у сигов. Подсчет числа тычинок ведется на левой первой жаберной дужке, иногда полез но учитывать отдельно число тычинок на верхней и нижней частях жаберной дужки; при исследовании рыб малого размера необходимо пользоваться бинокуляром или лупой. Просчитываются все тычинки, в том числе и зачаточные, которые бывают плохо видны, особенно у многотычинковых форм сигa и других видов.
Некоторые авторы отмечали увеличение числа тычинок с возрастом рыб, часто это относится к видам с большим числом тычинок типа ряпушки, пеляди, омуля [Букирев, 1938; Есипов, 1938; Меньшиков, 1951; Покровский, 1953; Канеп, 1971]. Но следует иметь в виду, что иногда такие выводы делаются на ма лых выборках (нет статистически достоверных различий), а, кроме того, у молодых рыб при большом числе жаберных тычинок легко пропустить зачаточные. Поэтому авторы, работающие с большими выборками, склонны считать, что у ряпушки [Гальцова, 1953; Лопатышкина, 1957; Никаноров, 1964), байкальского ому ля [Смирнов, 1974; Смирнов, Шумилов, 1974), сигa [Яковлева, 1968; Jarvi, 1940] число тычинок не меняется с возрастом. В про тивоположность этому у нельмы отмечается уменьшение числа тычинок с возрастом [Меньшиков, 1935; Красикова, 1960], что
Таблица 1. Возрастные изменения меристических признаков чунозерского сина (по сборам 1961-1971 гг.) (указаны пределы колебаний, средняя, ошибка средней, коэффициент вариации, число рыб)
Число жаберных тычинок 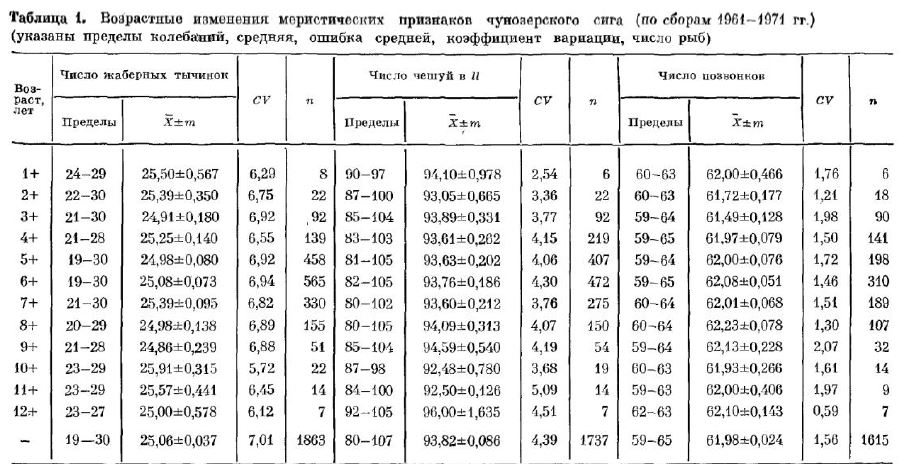
Таблица 2. Меристические признаки чунозерского сигa по сборам разных лет (пределы колебаний, средняя, ошибка) 

можно объяснить или редукцией крайних тычинок в связи с пере ходом к хищному образу жизни, или селективной смертностью особей с разным числом Тычинок. Небольшое снижение числа жаберных тычинок с возрастом отмечалось у хищного озерного Тольца из водоемов Камчатки [Решетников, 1961; Савваитова,
Решетников, 1961].
учунозерского сига окончательное число тычинок устанав ливается на первом году жизни и не меняется в онтогенезе (см. табл. 1). Постоянное число тычинок в пробах рыб разного воз раста отмечалось нами уанадырских сигов (востряк и горбун), чира и сибирской ряпушки.
На примере сигa с. lavаrеtus доказано, что число жаберных Тычинок контролируется генетическим фактором; этот признак передается по наследству от родителей потомству и в меньшей степени зависит от условий жизни. Проиллюстрируем это поло
жение несколькими примерами.
В потомстве сигов всегда четко наследуется мода по числу жаберных тычинок, поэтому даже при пересадке в новый водоем отклонения среднего числа жаберных тычинок невелики (рис. 5). При скрещивании самцов и самок с одинаковым числом Тычинок (брались крайние варианты для данной популяции) в потомстве хотя и наблюдаются колебания числа тычинок, но средняя близка к исходному числу тычинок у родительских пар (рис. 6). При скрещивании разных форм сига (мало- и многотычинковая) получается потомство с промежуточным числом тычинок, этот новый признак передается по наследству достаточно долгое время (рис. 7).
Опыты по акклиматизации двух форм сигa (лудога и чудской) в Севане показали, что в природных условиях наряду с новой гиб ридной формой довольно долго сохраняются родительские черты, Эти данные представлены на рис. 8, который составлен по мате риалам И. Ф. Правдина [1925, 1931], Р. С. Маиляна [1954, 1957),
Г. Х. Шапошниковой [1973), А. А. Никитина [ 1976] с добав
лением данных С. А. Пивазяна Число позвонков
за 1971 — 1972 и 1975— 1976 гг.
(который любезно предоставил Пределы
их в наше распоряжение) и на
ших сборов севанских сигов (1967 г.) и чудского сига (1975— 1976). Даже, при пересадке новой гибридной формы севанских сигов в оз. Иссык Куль число тычинок у потомства в новых условиях повторяет Число тычинок родительского севанского стада: в 03. Иссык Куль — 21 — 37, в среднем 30, 46, ав малом озерке Восточного Пирииссыккумья — 20— 38, в среднем 30, 85 жаберных тычинок.
В то же время неизвестно ни одного случая быстрого превра щения многотычинкового сигa в малотычинкового или наоборот [Берг, 1915]. Многовековая история сигов из альпийских озер служит тому примером [Thienemann, 1911, 1914; Surbeck, 1921; Kreitmann, 1924, 1929; Wagler, 1933; Dottrens, Quartier, 1949; Dot trens, 1950, 1955, 1959; Steinmann, 1950а, ъ, 1951; Batias, 1958; Spillmann, 1958; Vivier, 1958; Berg, Grimaldi, 1965; Lampert, 1971; Spreafico et al., 1974]. Закономерные изменения числа тычинок сигов обычно связаны с характером питания. Так, Ярви [Jarvi, 1928] приводит данные по увеличению числа тычинок у более юж ных популяций балтийского сига, что связано не только с темпе pатуpoil развития этих рыб, но и с обильным развитием планктона в более южных районах. Аналогичное явление отмечено и для си гов Северной Америки: северные популяции американских сигов имеют меньшее число жаберных тычинок по сравнению с южнымII (Vladykov, 1934; Dymond, 1943; Lindsey, 1926b; Lindsey et al., 1970). Если нанести на карту среднее число тычинок да я разных популяций, то получается наглядный пример клинальной изменчивости признака. Такие карты приводятся для сигов Балтийского бассейна (Himberg, 1970] и сигов Северной Америки [Lindsey et al., 1970]. В гл. VI приводятся такие данные для сигов Сибири и пограничных районов (см. рис. 45). В табл. З приведены данные по тычинкам сигов Кольского полуострова.
Распределение популяций с разным числом тычинок в Европе имеет определенную широтную зональность. Ареал многотычинковых сигов приурочен к Балтийскому бассейну. Так, в реках и озерах Северной Лапландии (Северная и Швеция, Северная Финляндия, Кольский полуостров) имеется четкая бимодальная кривая частоты встречаемости популяций с разным числом тычинок, которая соответствует двум формам снигов: малотычинковой с 20-30 Тычинками с бентосным питанием и многоТычинковой (сиг 
Рис. 5. Частоты встречаемости сигов с разным числом жаберных тычинок при выращивании особей одной популяции в разных условиях обитания [10; Svardson, 1957, 1970) 1 — родительское стадо, р. Индастьсельвен (455 экз.); ии -- сиги, выращенные в бо лее теплом пруду Калаpнe (38 экз.); иии — сиги, аклиматизированные в 03. Коль сон, спустя 10 лет после пересадки из р. Индaльсельвен (602 экз.); IV — эти же сиги сатустя 20 лет после пересадки (602 экз.)
Рne. 6. Распределение числа особей с разным чис,0м жаберных тычинок среди родительского стада и потомства, полученното при скрещивании пар производителей с крайними вариантами числа тычинок [по: Svardson, 1970] 1 — родительское стадо сигов, o3. идсьон (782 экз.); ии — потомство от родителей с 32х32 тычинками (120 экз.); иии — потомство от родителей с 41?41 тычинками (40 экз.)
планктофаги) с 30 — 50 тычинками. В водоемах средней Финлян дии и Карелии появляется в большом количестве промежуточная -
форма (возможно, гибридного происхождения) и кривая уже не имеет четких двух вершин. Эти формы сигов прослеживаются до альпийских озер (рис. 9).
Таким образом, подмеченная закономерность изменения числа жаберных тычинок в зависимости от широты расположения водоема, с одной стороны, связана с разными генетическими линия ми сигов в водоемах балтийского бассейна (мало- и многотычин ковые формы), а с другой стороны, отражает характер питания и преобладание в питании южных популяций более мелких кормо вых организмов [Walczak, 1953; Svardson, 1952]. Аналогичным образом можно объяснить уменьшение числа жаберных тычинок у глубоководных форм в связи с переходом на более крупные кор ма [Koell, 1931].  — скрещиваемые родительские формы: самки малотычинкового сигa (17 экз.) и самцы многотычинкового сига")
Рис. 7. Число жаберных тычинок у двух исходных родительских форм сигa и трех поколений гибридов [по: Svard son, 1970) — скрещиваемые родительские формы: самки малотычинкового сигa (17 экз.) и самцы многотычинкового сига (26 экз.); II — гибриды первого поколения (528 экз.); III — гибриды второго поколе ния (430 экз.); IV - гибриды третьего поколения (586 экз.)


Между числом жаберных тычинок на первой дуге и относи тельной длиной наибольшей якаберной тычинки имеется прямая коррелятивная связь, что можно продемонстрировать на примере -сигов из лапландских озер [Решетников, 1963ail: 
Канадские авторы [Scott, Smith, 19621 предлагают для разде ления ряпушек оз. Эри пользоваться суммой числа тычинок и от носительной длины центральной ?аберной тычинки (в % к длине жаберной дуги), полагая, что по этому показателю можно разгра ничить два вида: C. alpenae имеет условный индекс 64-77, в среднем 69,2; C. artеdии — 82— 96, в среднем 89,5.
Иногда для практических целей пытаются строить рабочую схему для разделения форм сигов только по числу жаберных ты чинок и размерам тела [Китаев, 1976]. Как ни велика таксономи ческая значимость числа жаберных тычинок, нельзя построить систему сигов только по одному этому признаку; примером тому является искусственная система Г. Свердсона [Svardson, 1957а).
Число пилорических придатков относится к наиболее вариа бельным из всех меристических признаков у рыб. Оно находится в тесной зависимости и от характера питания данной популяции или вида, что было показано на примере сельдевых, тресковых, лососевых, осетровых, камбаловых, бычковых и других рыб | Све товидов, 1932, 1953; Решетников, 1961; Каневская и др., 1968; Савваитова, 1976; Vladykov, 1934; и др.]. Число пилорических придатков цих длина меняются с возрастом, как правило, больше, чем у хищных рыб. Таксономическое значение этого признака не велико, но полагают, что он может служить видовым критерием для некоторых сиговых рыб [Vladykov, 1934; Himberg, 1970].
В табл. 4 приведены данные о числе жаберных тычинок, чешуй в боковой линии, позвонков и пилорических придатков у сиговых из водоемов СССР. Эти данные несколько отличаются от тех, что приводились нами ранее (Nikolsky, Reshetnikov, 1970], поскольку они дополнены нашими сборами по нельме, чиру, сигу, ряпушке и вальку и многочисленными новыми материалами, опубликованными в последние годы. Данные из литературы сопоставлены между собой, сверены с нашими данными и музейными коллек циями; число позвонков проверено по рентгеноснимкам коллекций ЗИН АН СССР. Все сомнительные и резко отклоняющиеся данные в табл. 4 не включены, в большой мере это касается числа позвон ков (более подробно об этом см. в гл. VI). Поэтому приводимые
в табл. 4 данные, с нашей точки зрения, характеризуют пределы с колебаний данного признака для вида в целом. Имеются лишь
небольшие исключения для некоторых зарубежных водоемов. Так,
для ряпушкин польских озер пределы колебаний числа пилориче в ских придатков равны 51 - 110 [Walczak, 1953]. У вида c. sardi
nella в водоемах Аляски и Канады число придатков имеет преде
лы 74—111 [McPhail, Lindsey, 1970), а для вида c. clupeaformis - вместе с пыжьяном отмечают от 140 до 264 пилорических придат
ков [Koelz, 1930; Dymond, 1943; McPhail, Lindsey, 1970; Scott, - Crossman, 1974].  и чудской сиг")
Рис. 8. Число жаберных тычинок у исходных родительских форму и смешан дой популяции севанских сигов 4 - сиг-лудога (123 экз.) и чудской сиг {80 экз.) в своих родных водоемах; ии – ce аванские сиги в 1950-х годах (200 экз.); иии — севанские сиги в 1967—1971 п.
491 экз.); IV — севанские сигун в 1975—1976 гг. (237 ?кз.) 
Рис. 9. Среднее число жа берных тычинок у популяций сигa из водоемов Ceверной Швеции и Лапландии (I), Средней Финлян дии и Карелии (II) и Альпийской зоны ( по: Himberg, 1970, с добавлением наших данных
По степени изменчивости меристических признаков все исследо ванные популяции сиговых рыб в порядке возрастания коэффициентов вариации (си) располагаются в такой последовательно сти: позвонки — число чешуй в 21 — ветвистые лучи в P, V, D, А — жаберные тычинки —— пилорические придатки (табл. 5). Мы полагаем, что степень изменчивости признака зависит от сроков, когда формируется окончательное число счетных элементов в он тогенезе: чем раньше закладывается признак, тем меньше его степень изменчивости. Возможно, это является специфичным не только для сиговых рыб, но и для многих других, поскольку ана логичная закономерность подмечена нами для лососевых, кардо вых и окуневых рыб. Примерно такую же последовательность по степени изменчивости меристических признаков отмечал С. В. Канеш [ 1976] Для ряпушки, рипуса, сибирской ряпушки, пеляди, тугуна, омуля и муксуна; он долагал, что такая последо вательность признаков по степени их изменчивости «свидетель ствует о порядке их наследуемости» [Канец, 1976, с. 618]. Это не совсем верно, скорее эта последовательность отражает продолжи тельность влияния внешних условий на формирование окончательного числа счетных элементов в онтогенезе.
Анализ наших данных показывает, что коэффициент вариации зависит от значения измеряемого признака, на что обращал вни мание еще Э. Майр -[ 1971]. Обычно коэффициент вариации боль
Таблица 4. Пределы колебаний некоторых меристических признаков сиговых рыб из водоемов СССР 
шинства пластических и мерчистических признаков не превышает 10%, что свидетельствует об однородности выборок и небольших погрешностях промеров. Однако коэффициенты вариации верхне челюстной кости (10— 12% ), высоты рыла (12%) и других малых величии бывают явно завышены (до 28%). Причины этого явле ния имеют двойственный характер. С одной стороны, это отражает большую изменчивость самого признака: у многих сиговых рыб на верхней челюсти имеется хрящевої вырост, поэтому трудно бывает замерить с достаточної точностью высоту и ширину рыла, - длину верхнечелюстной кости и некоторых других. С другой сто роны, коэффициент вариации, как выражение среднеквадратич ного отклонения в процентах от средней, зависит от значения приз нака: чем меньше значение признака, тем выше коэффициент ва Триации и наоборот. Особенно наглядно это проявляется при самых крайних значениях средних величин (рис. 10). По этой причине 3 табл. эне включено число неветвистых лучеиї в плавниках, так как при средних порядка 3,5-4,0 и небольших средних квадра яичных отклонениях порядка 0,3—0,6 коэффициенты вариации - равны 13—21 %.
.. Таксономическая значимость ме истических признаков. Таким образом, приведенные выше материалы позволяют сделать сле ?дующее заключение. Меристические признаки можно использо вать в таксономических II популяционных исследованиях, если сборы охватывают рыб разного возраста и выборки достаточно велики (не менее 30 экз.) или если пробы собирались в течение
Таблица 5. Коэффициенты вариации меристических признаков у исследованных популяций сиговых рыб 
 и меристических признаков (2) и их коэффициента и вариации на примере анадырcкого валька")
Рис. 10. Соотношение между значениями пластических (1) и меристических признаков (2) и их коэффициента и вариации на примере анадырcкого валька [Решетников и др., 1975]
ряда нет. В этом случае возможные небольшие различия средних значенийї у рыб разных поколений (генераций) нивелируются. Можно полагать, что каждую популяцию характеризует своїй устойчивый набор меристических признаков, что !!озволяет исполь зовать их для внутривидовой систематики. Из всех счетных приз наков наиболее ценным является число жаберных тычинок, так как у синговых рыб число тычинок находится под генетическим контролем II их можно использовать в качестве «генетического маркера» [Koelz, 1930; Dymond, 1943; Svardson, 1952, 1970; Lind sey et al., 1970].