2. Репродуктивные признаки самок
Исследование репродуктивных показателей производителей имеет важное значение и для рыбоводных, и для селекционных работ, поскольку эти признаки в значительной мере определяют продуктивные качества маточных стад рыб. К ним относятся плодовитость, качество икры, выживаемость потомства. Непосредственное отношение к оценке воспроизводительной способности самок при искусственном воспроизводстве имеет также срок созревания рыб в течение нерестового сезона.
Срок созревания в нерестовом сезоне. Нерест в популяциях самых различных видов рыб с единовременным икрометанием представляет собой длительный процесс, растягивающийся иногда на несколько месяцев. В этом отношении не составляют исключения и сиговые рыбы, в частности, пелядь. По данным Е.В. Бурмакина (1953) и Б. К. Москаленко (1955), продолжительность нерестового сезона у пеляди в сибирских реках достигает 1-16 месяцев, а в оз. Ендырь – 30-40 дней (Никонов, 1963; Кугаевская, 1978). Как и у других видов рыб, продолжительность нереста у пеляди определяется многочисленными причинами. По-видимому, в первую очередь она зависит от комплекса гидрометеорологических условий, из которых решающая роль принадлежит температурному фактору, причем не только температурным пределам, но и характеру термического режима водоема в нерестовый период. Кроме того, на длительность нерестового сезона может оказывать влияние возрастной и размерный состав рыб, их физиологическое состояние и т. п. (Терещенко, 1913, 1917; Черфас, 1940; Анохина, 1969).
Как показывает рыбоводная практика, растянутость нереста во времени сохраняется и в условиях разведения – при использовании для Получения потомства производителей, выращенных в прудах и озерах. В этих условиях значительно легче исследовать роль отдельных факторов, определяющих длительность нерестового периода, уравняв влияние остальных: например, изучая стадо одновозрастных производителей, выращенных в одинаковых условиях. Такое исследование проводилось с растительноядными рыбами и показало, что разновременность созревания самок белого толстолобика и белого амура ежегодно проявляется в одновозрастном маточном стаде, выращенном в условиях рыбоводного хозяйства вне ареала (Слуцкий, 1971д; Андрияшева, 1973б). Часть этих самок была исследована гистологически, и выяснилось, что процесс вителлогенеза проходит у них асинхронно, приводя к разновременному созреванию самок белого толстолобика в течение нерестового сезона (Сарсембаев, 1972,1974). Позднее гетерогенность одновозрастных самок по сроку нереста была обнаружена у пестрого толстолобика, акклиматизированного в Средней Азии (Сарсембаев, 1978). Аналогичных сведений о сиговых рыбах в литературе мы не обнаружили. Известно только, что в маточном стаде пеляди разного возраста существует асинхронность созревания особей, обусловленная тем, что начало трофоплазматического роста ооцитов растянуто у них на 25-30 дней (Кузьмин, 1967,1975).
С целью исследования разнообразия самок ендырcкoй пеляди по сроку нереста в течение 1973-1979 гг. ежегодно определяли: 1) продолжительность нерестового сезона для самок одного возраста, 2) распределение самок в течение этого периода (т. е. количество созревших рыб из числа просматриваемых каждые 3-4 дня) и 3) индивидуальный срок нереста для каждой самки.
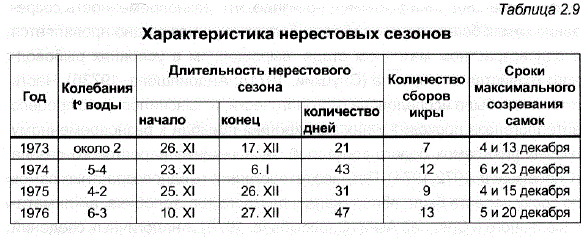
В период с 1973 по 1976 г. было изучено 438 самок из первого ендырского стада. Полученные за четыре нерестовых сезона данные (табл. 2.9) могут свидетельствовать о том, что у eндырcкoй пеляди в условиях искусственного воспроизводства наблюдается значительная гетерогенность по сроку созревания самок в нерестовом сезоне.
В зависимости от температурных условий сезона растянутость нереста проявляется в большей или меньшей степени: при более низкой температуре воды нерест проходит в более сжатые сроки – 20-30 дней (1973 и 1975 гг.), а при более теплой осени растягивается до 1,5 мес. Тем не менее независимо от температурных условий сезона созревание самок в каждом из них имеет два максимума. Бимодальность кривой распределения становится более наглядной при обобщении данных, полученных за 4 нерестовых сезона (рис. 2.4). Необходимо подчеркнуть при этом, что выявленная гетерогенность, характеризующая рыб одного возраста, сходного размера, веса и физиологического состояния, не меньше той, которая наблюдается в нерестовых стадах в природных популяциях. населени менен
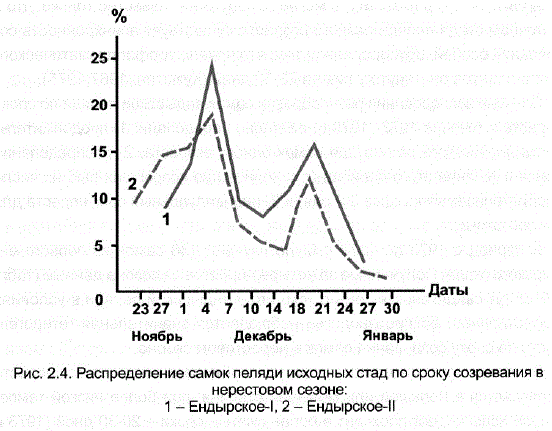
Наиболее важным из экспериментальных данных является наличие на кривой распределения самок по времени созревания двух модальных классов, которые были обнаружены в четырех нерестовых сезонах. Этот факт, по-видимому, может свидетельствовать о существовании в стаде двух групп производителей, качественно различающихся по изучаемому признаку, в данном случае – по времени нереста. Мы условно назвали их нерестовыми группами: І нерестовая группа объединяет раносозревающих в сезоне самок, II — поздносозревающих.
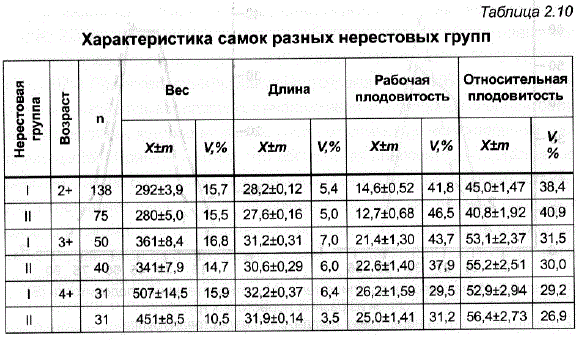
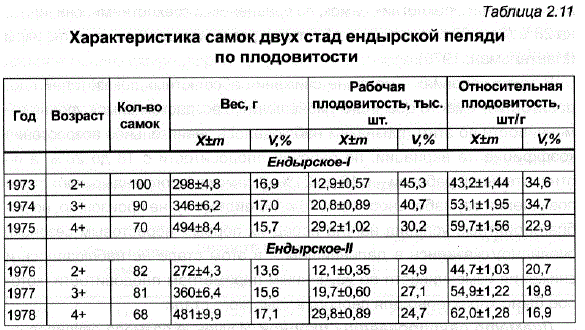
При сравнении самок из разных нерестовых групп по нескольким рыбоводно-биологическим показателям (табл. 2.10) удалось выявить небольшую разницу между ними по длине тела и по весу. У трехлеток она была достоверна (р<0,001 и р<0,05 соответственно), у четырехлетних рыб не достигала статистически значимых величин, а у пятилеток оказалась существенной только по весу (р<0,001). При первом нересте раносозревающие самки были более плодовиты, чем поздносозревающие (р<0,05).
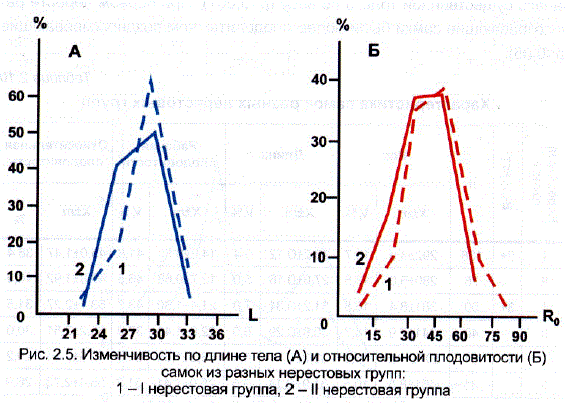
В среднем же за три года исследований между этими группами рыб не удалось обнаружить существенных различий по рабочей и относительной плодовитости. Вместе с тем разделение стада на две группы по времени нереста не привело к отклонению групповых кривых распределения самок по длине и относительной плодовитости от нормальных (рис. 2.5).
Коэффициенты изменчивости внутри каждой из групп не отличались от средних для этого маточного стада. Ногоон наполовина на Bн
Тенденция к снижению некоторых рыбоводно-биологических показателей производителей на протяжении нерестового сезона наблюдалась и при исследовании некоторых других объектов рыбоводства. Так, найбольшее уменьшение длины и плодовитости самок в конце нерестового периода отмечено у белого амура Е.С. Слуцким (1971д). Более крупными и плодовитыми оказались раносозревающие самки белого толстолобика (Андрияшева, 1973б) и форели (Щербенок и др., 1982). На наш взгляд эти факты, как и данные по пеляди, свидетельствуют не о качественных различиях между группами рыб, различающихся по сроку нереста, а вероятно, об определенных особенностях созревания, свойственных самкам одной популяции независимо от видовой принадлежности.
Дополнительное доказательство подразделенности самок ендырской популяции пеляди по сроку созревания было получено при исследовании второго маточного стада ендырского происхождения (завоз икрой в 1973 г.). На рис. 2.4 видно, что суммарная кривая распределения частот для самок || eндырского стада (n=436 шт.) за три нерестовых сезона (1976–1978 гг.) практически совпадала с аналогичной кривой для самок
Тендырского стада. Бимодальность кривой второго стада сохранялась все три года так же устойчиво, как и первого. Несмотря на различия в температурных условиях нерестовых сезонов в период с 1973-1978 гг., усредненные максимумы на обеих кривых соответствовали одним и тем же датам — 4 и 18 декабря. а в совокупности все эти факты свидетельствовали о наличии определенной, устойчиво сохраняющейся во времени при искусственном воспроизводстве популяционной структуры в исследованных стадах ендырской пеляди. Как выяснилось позднее (Кугаевская, 1978), наши данные полностью совпали с теми, которые были получены при изучении нерестовой Популяции в маточном водоеме: в оз. Ендырь пелядь нерестилась 40 дней, и 2 максимума на кривой распределения самок V стадии зрелости приходились на 4-еи 16-е декабря. Таким образом, стало очевидным, что обнаруженная подразделенность самок исследованных маточных стад является отражением специфических особенностей популяционной структуры пеляди из оз. Ендырь.
Плодовитость. Изучению плодовитости рыб посвящено огромное количество ихтиологических исследований, объектом которых обычно является величина абсолютной плодовитости (Суворов, 1948; Анохина, 1969; Никольский, 1974). При проведении рыбоводных и селекционных работ изучение плодовитости сводится, как правило, канализу величины рабочей плодовитости, Количеству икры, которое можно получить от самки при Искусственном воспроизводстве. Принято считать, что абсолютная плодовитость всегда выше рабочей. Так, По сообщению П. А. Дрягина (1948), рабочая плодовитость пеляди составляет лишь 73% от абсолютной. Однако, как показал наш опыт работы с сигами, при своевременном использовании самок для получения икры и правильном ее отцеживании величина Индивидуальной абсолютной и Индивидуальной рабочей плодовитости практически совпадает.
Сведения о плодовитости ендырcкoй пеляди в литературе немногочисленны и касаются только абсолютной и средней рабочей плодовитости. Величина абсолютной плодовитости пеляди в оз. Ендырь в среднем составляет 14-16 тыс. икринок, при колебании от 8 до 28 тыс. (Москаленко, 1955); позднее было показано, что она может достигать 46,9 тыс. шт. (Никонов, 1963; Москаленко, 1971). При выращивании производителей ендырcкoй пеляди в прудах Украины абсолютная плодовитость 2-5-летних самок равнялась 21,5 тыс. ШТ. при колебании от 9 тыс. у двухлеток до 40 тыс. у пятилеток (Носаль, Менюк, 1958; Носаль, 1968). Г.А. Головков и А.Н. Кузьмин (1963) полагают, что от трехлетней самки весом 450-500 гможно получить 25 тыс. икринок; расчетная плодовитость пеляди весом 700-800 г принимается равной 30 тыс. шт. (Головков, Кузьмин, 19
Однако обработанные нами данные Г.П. Андрушайтис, приведенные 3 Г. А. Головковым и А. Н. Кузьминым (1963), свидетельствуют о том, что на рыбоводном заводе Томе средняя рабочая плодовитость не превышала 15 тыс. икринок (по 77 самцам). В Ропше, где в нерестовую кампанию используют 2-2,5 тыс. самок в основном 3-5-летнего возраста, в среднем от самки получали 20,9 тыс. икринок (по данным ЦЭС «Ропша», 1970–1979 гг.).
Для селекции, в отличие от рыбоводства, наибольший интерес представляют индивидуальные показатели плодовитости и их изменчивость, а не средняя величина рабочей плодовитости, рассчитываемая на одну самку и неверно трактуемая в рыбоводной литературе как индивидуальная рабочая плодовитость (Анохина, 1969). Для оценки исходного уровня разнообразия в маточном стаде ендырcкoй пеляди были использованы индивидуальные характеристики самок по двум параметрам плодовитости – рабочей и относительной рабочей (отношение величины рабочей плодовитости к весу тела самки без икры), что позволяло получить более полное представление о воспроизводительной способности различных самок (ЗоНова, 1976; Спановская, 1976). в Как показали наши данные, неблагоприятные условия выращивания маточного стада, отличавшиеся особенно напряженным кормовым режимом в 1974 и 1976 гг., привели не только к отставанию в росте, но и были причиной невысокой плодовитости (в среднем около 13 тыс. шт.) у впервые созревших в прудах Ропши трехлеток ендырcкoй пеляди (см. табл. 2.10). В дальнейшем наблюдалось увеличение рабочей и относительной плодовитости с возрастом: по рабочей плодовитости 5-летние самки преВосходили трехлеток в 2,3 раза, по относительной – в 1,4 раза. В итоге от пятилетних самок весом 500 г было получено 29-30 тыс. икринок.
Коэффициенты изменчивости исследованных параметров плодовитости свидетельствовали о том, что они являются наиболее варьирующими у самок признаками. Максимальная изменчивость была обнаружена у впервые нерестящихся производителей: рабочая плодовитость трехлетних самок, выращенных в одном пруду, различалась более чем в 6 раз, а относительная – в 4 раза. Такие результаты в значительной степени были обусловлены наличием в первом стаде некоторого количества самок (около 10%), отдавших небольшое количество икры. Это могло быть Связано не только с тем, что в первый нерестовый сезон трехлетки могли иметь низкую абсолютную плодовитость, но ис несовершенством биотехники сбора икры — чаще всего преждевременным отцеживанием икры, а поэтому неполным. При ликвидации этих недостатков процент самок с минимальным количеством икры значительно сократился, и коэффициенты изменчивости по этому признаку у трехлеток второго ендырского стада уменьшились до 25-27% (табл. 2.11) В большинстве случаев кривые изменчивости по рабочей и относительной плодовитости подчинялись закону нормального распределения и характеризовались умеренной концентрацией вариант вокруг средней.
С увеличением возраста самок изменчивость по исследованным параметрам плодовитости снизилась и стабилизировалась на уровне 20-30%. Аналогичные результаты были получены при изучении индивидуальной плодовитости у других объектов рыборазведения. Так же, как и у пеляди, этот признак отличался особенно высокой вариабельностью у самых различных видов рыб: у чудского сигa коэффициенты изменчивости составляли 37-42% (Тищенко, 1978а), у радужной форели — 25-40% (Gall, 1975; Щербакова, 1981; Щербенок и др., 1982), у атлантического лосося – 30-36% (Казаков, Мельникова, 1980), у белого толстолобика – 28-30% (Андрияшева, 1973а) и достигали в некоторых случаях у белого амура 50% (Слуцкий, 1971д, 1973а).
Согласно многочисленным ихтиологическим и рыбоводным исследованиям, величина плодовитости рыб в значительной степени зависит от условий нагула маточного стада и, в частности, от обеспеченности рыб кормом (см., например, Никольский, 1953, 1965; Анохина, 1969; Зонова, 1976; Крохалевский, 1980).
Нам предоставилась возможность исследовать величину плодовитости и ее изменчивость у пеляди при чрезвычайно неблагоприятных условиях, которые сложились в нагульном пруду в 1976 г. из-за необычайно холодного лета и 3-кратной плотности посадки рыб. В результате существенного недостатка зоопланктона величина относительного прироста самок по весу за это лето оказалась отрицательной (-13%), рабочая плодовитость четырехлетних самок, по сравнению с трехлетними, снизилась на 28% (2+ – 20 тыс. шт., 3+ — 15,5 тыс.), резко ухудшилось качество икры (Мантельман, 1976).
В таких условиях – на фоне снижения абсолютных показателей плодовитости – вместо обычного уменьшения с возрастом самок уровня изменчивости по этим признакам наблюдалось значительное возрастание коэффициента вариации: по рабочей плодовитости с 18 до 28%, а по относительной рабочей — с 16 до 21%. У самок второго ендырского стада Повышения вариабельности в 1976 г., по-видимому, не произошло, но неблагоприятные условия нагула перед их первым нерестовым сезоном, очевидно, сказались в дальнейшем — в этом стаде не наблюдалось и снижения изменчивости по изученным параметрам плодовитости, которое было характерно для первого ендырского стада.
Диаметр овулировавших икринок. Изучение размера овулировавших икринок является важнейшей составной частью исследования воспроизводительных способностей самок, так как по этому показателю и его изменчивости можно судить о качестве икры. Поэтому неудивителен тот большой интерес, который с давних пор проявляют к этому признаку ихтиологи и рыбоводы (см. Слуцкий, 1980). Согласно сложившемуся в ихтиологии представлению, больший размер икринки означает лучшее снабжение эмбриона питательными веществами (Терещенко, 1917; Шмальгаузен, 1935; Мейен, 1940; Суворов, 1948; Матвеев, 1951; Смирнов, 1954; Семенов, 1963; Волкова, 1965 и др.). У самых разных видов рыб размеры икринок оказывают существенное влияние на многие свойства личинок и даже молоди, определяя их величину, выживаемость, степень развития (Морозов, 1951; Garside, Fry, 1959; Ястребков, 1966; Песлак, 1968; Fowler, 1972; Gall, 1974; Зонова, 1974а; Яндовская, 1976; КазаКов, Мельникова, 1980; Рязанцева и др., 1981). По мнению Е.С. Слуцкого (1980) и Р. В. Казакова (1982), уровень индивидуальной изменчивости размеров овулировавших икринок является важным показателем физиологического состояния самок и может служить надежным критерием для их оценки при проведении рыбоводных и селекционных работ.
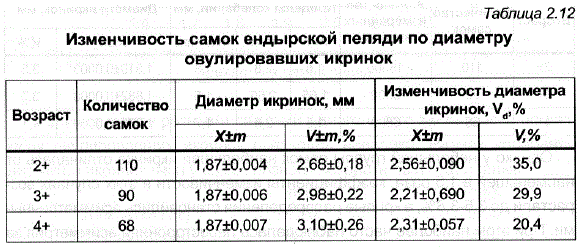
Как показало исследование размеров икры у самок первого ендырского стада (1973–1975 гг.), средний диаметр овулировавших икринок у трехлетних самок пеляди был равен 1,81 мм, у четырехлеток он увеличился до 1,87 мм (р<0,001), у пятилетних больше не изменялся (табл. 2.12). Увеличение размера икринок с возрастом самок известно для многих видов рыб, в том числе и для объектов искусственного разведения — белого амура, карпа, форели, атлантического лосося, чудского сига (Buss, McCreary, 1960; Галкина, 1970; Слуцкий, 1971а; Зонова, 1973; Gall, 1974; Савостьянова, Слуцкий, 1974; Тищенко, 1976б; Мельникова, Лейзерович, 1979; Казаков, Мельникова, 1980,1981).
У пеляди, как и у других рыб, особенности индивидуального развития икринок привели к разноразмерности овулирующих у одной самки икринок. При этом оказалось, что у разных самок пеляди икринки в разной степени различались между собой по диаметру. У большинства самок отношение max/min составляло 1,0-1,2, коэффициенты вариации колебались у них от 1,2-1,5 до 3,0%, кривые изменчивости были симметричны (рис. 2.6).
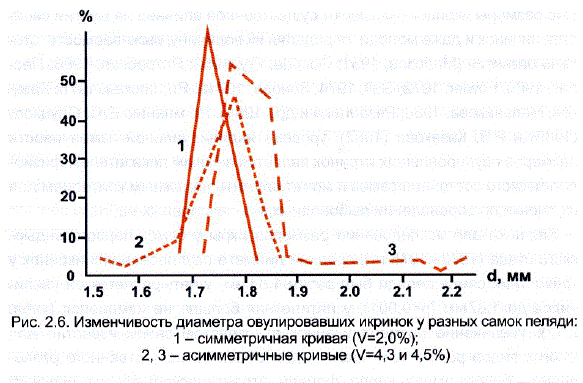
Однако у небольшой группы самок наибольшая икринка отличалась от найменьшей в 1,5 раза, коэффициенты изменчивости в этих случаях возрастали до 3,5-3,8%, а кривые распределения становились асимметричными. При этом наиболее часто наблюдалась левосторонняя асимметрия за Счет наличия в пробе нескольких крупных набухших икринок (см. рис. 2.6). Увеличение левого плеча распределения было отмечено за три года только у нескольких самок. В целом такие различия можно было наблюдать у самок всех возрастов, хотя в среднем, как это видно из табл. 2.12, степень «внутрииндивидуальной» вариабельности с увеличением возраста самок Снижалась с 2,56 до 2,31%. Аналогичная закономерность была обнаружена у карпа (Зонова, 1973) и у форели (Савостьянова, Слуцкий, 1974).
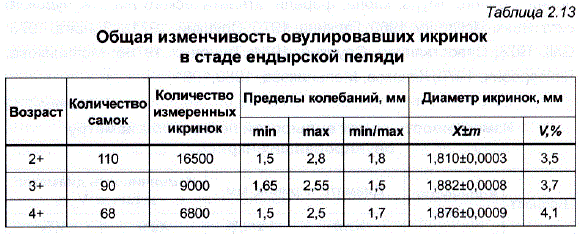
Суммирование данных, полученных при измерении всех икринок у самок одного возраста, дает представление о величине общей изменчивости икринок по диаметру (табл. 2.13).
Как и у других рыб, у пеляди диапазон индивидуальной изменчивости икринок для популяции в целом значительно больше – наименьшие икринки отличались от наибольших на 43-45%, а коэффициенты вариации возрастали до 3,5-4,0%. Кривая изменчивости соответствовала закону нормального распределения (рис. 2.7).
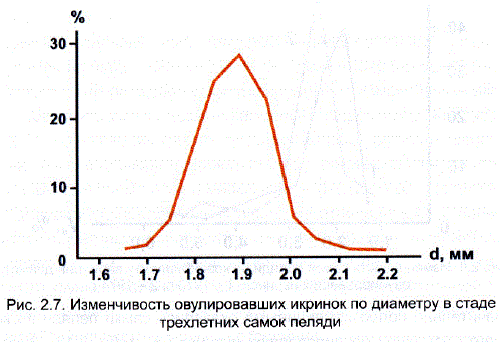
Различия между разными самками одного и того же возраста по размеру овулировавших икринок оказались невелики. Размах колебаний диаметра икринок (мм) у самок разного возраста был стабильным:
Таким образом, самки, имеющие наиболее крупную икру, отличались от самок с мелкой икрой на 15-7%. Изменчивость самок по среднему диаметру икринок была низкой и практически не менялась с возрастом (см. табл. 2.12), что может свидетельствовать о значительно большей стабильности этого признака у пеляди по сравнению с растительноядными, карпом, форелью, чудским сигом, и лососем (Слуцкий, 1971а, б; Зонова, 1973; Савостьянова, Слуцкий, 1974; Тищенко, 1976б; Мельникова,, Лейзерович, 1979). - в большей степени отличались разные самки пеляди по такому показателю, как количество овулировавших икринок, содержащихся в 1 г. Средняя величина этого показателя у четырехлетних самок составила 291+2,7 икринок, а коэффициент изменчивости был равен 7,9%.
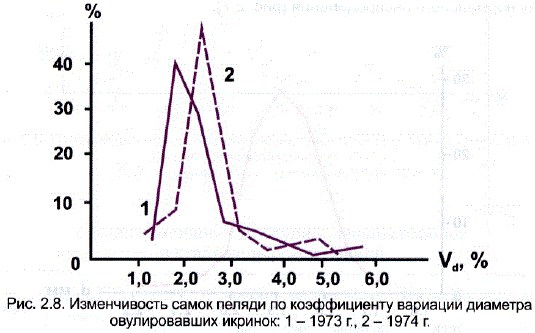
Значительно более изменчивыми оказались самки пеляди по коэффициенту вариации овулировавших икринок (см. табл. 2.12). Значения средних по этому показателю, как уже отмечалось, очень малы и свидетельствуют о низком уровне индивидуальной изменчивости икринок у каждой самки. По мере роста самок они еще уменьшаются. Изменчивость самок по коэффициенту вариации диаметра икринок, т.е. коэффициенты вариации второго порядка (V.%), резко увеличена. Кривые изменчивости по этому показателю (рис. 2.8) имеют сильно вытянутое правое плечо за счет группы самок, отличающихся повышенной вариабельностью икры (V>3,0%). Таких самок среди исследованных в разные годы оказалось: в 1973 г. – 17, в 1974-м — 18 ив 1975-м — около 6%.
Увеличение изменчивости икринок по диаметру за счет появления у некоторых самок небольшого количества крупных, набухших икринок наблюдали при искусственном воспроизводстве сазана, белого амура, толстолобиков и обычно объясняли перезреванием икры из-за задержки использования этих самок для получения потомства (Слуцкий, 1971а, б, 1973б; Ненашев, Боржемская, 1974). По мнению И.И. Мантельман (1978), наличие среди самок пеляди особей, отличающихся повышенной вариабельностью икринок, также является следствием несвоевременного отцеживания у них икры. Как показало изучение размеров икринок у самок чудского сига, отловленных во время нерестового хода в Чудском озере (Тищенко, 1976б), только единичные особи имеют несколько повышенный коэффициент изменчивости, а вариабельность самок по этому показателю значительно ниже, чем у других видов рыб, разводимых искусственным путем.
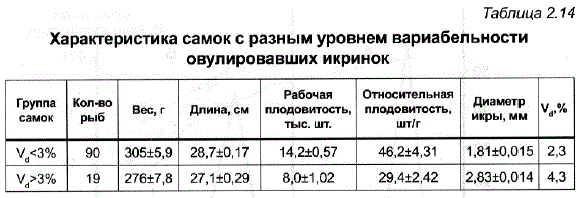
Однако исследование некоторых рыбоводно-биологических показателей у самок пеляди, различавшихся по степени вариабельности икринок, выявило, что повышенную индивидуальную изменчивость икринок нельзя объяснить только несвоевременным сбором икры. Как видно из табл. 2.14, такие самки отличались низкими продуктивными качествами в целом – меньшим весом и длиной (р<0,05), пониженной плодовитостью (р<0,01). В то же время группа мелких самок (I=27,310,20) с нормальным уровнем вариабельности икринок (2,2%) не отличалась от среднего по стаду по относительной плодовитости (43,0+0,19). Все это может свидетельствовать о том, что уровень изменчивости икринок у разных самок зависит от функционального состояния их воспроизводительной системы и при отклонении этого состояния от нормы может приводить к нарушению стабильности процесса созревания.
Изменение уровня вариабельности размеров икринок может наблюдаться не только при несвоевременном отцеживаний икры, но и при значительных отклонениях от нормы условий содержания производителей. Как было показано выше, существенная нехватка корма в течение лета 1976 г. Повлияла не только на рост рыб: она привела к снижению плодовитости, резкому ухудшению качества икры, а также отразилась на размерах овулировавших икринок и уровне их изменчивости. Зависимость размеров икринок от условий выращивания половозрелых рыб – хорошо известный в рыбоводной литературе факт (см. Зонова, 1973; Слуцкий, 1980), гораздо менее изучены пределы изменения вариабельности этого признака.
По нашим данным, величина изменчивости самок пеляди по среднему диаметру икринок, обычно не изменяющаяся с возрастом, была минимальной при оптимальных условиях выращивания 1975 г. (V=2,24%) и существенно возросла (V=2,76%) при их резком ухудшении в 1976 г. (р<0,01). Индивидуальная изменчивость икринок у каждой самки, наоборот, резко снизилась (с 2,67 до 2,01%) при нехватке корма в преднерестовый период 1976 г. и увеличилась до 2,45% на следующий год при значительном улучшении условий содержания стада (р<0,01).
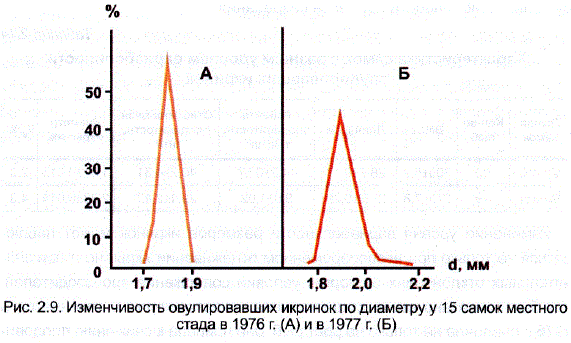
Как видно из рис. 2.9, где представлены кривые распределения икринок от 15 самок, имевших одинаковый модальный класс по диаметру (1,80 мм — в 1976 г. и 1,95 мм – в 1977 г.). В 1976 г. диапазон изменчивости был явно уже, чем в 1977 г. В первом случае (рис. 2.9, A) овулировавшие икринки распределялись по пяти классам и на модальный приходилось 58%, а во втором (рис. 2.9, Б) — по десяти классам, при снижении количества в моде до 42%. Следует особо подчеркнуть, что в наших опытах минимальный уровень «внутри- индивидуальной» изменчивости икринок был характерен для неблагоприятных условий.
Таким образом, крайние отклонения условий содержания самок от оптимальных могут приводить к нарушениям определенных закономерностей динамики изменчивости размера овулировавших икринок, которые были выявлены при исследовании пеляди первого ендырского стада и известны для некоторых других видов рыб (Слуцкий, 1980).