Наследуемость различных количественных признаков.
Биологические особенности рыб и своеобразие их популяционно-генетической структуры предоставляют большие возможности для корректного определения уровня генетической изменчивости по количественным признакам у большинства рыбоводных объектов. Следует, однако, признать, что реализация этих возможностей до недавнего времени вызывала серьезные затруднения. В связи с этим работы по изучению селекционно-генетических параметров в популяциях рыб стали появляться в литературе только в 70-х гг. Их стимулировала индустриализация рыборазведения, расширившая технические возможности для генетического анализа количественных признаков. За первые 10 лет этих исследований в монографиях и наиболее полных обзорных статьях были опубликованы данные по наследуемости многих хозяйственно важных признаков более чем по 15 видам рыб (Gjedrem, 1975, 1983; Кирпичников, 1979, 1983; Moav, 1979; Kirpichnikov, 1981). При этом у основных объектов разведения, таких как атлантический лосось и радужная форель, величина наследуемости была определена уже по 22-35 признакам. Из них более половины характеризовали уровень генетической изменчивости по скорости роста молоди разного возраста. Вместе с тем в 80-90-е гг. прошлого века это направление селекционных исследований в рыбоводстве значительно отставало от других отраслей селекции животных, и не только у нас в стране (несколько работ по карпу – Ненашев, 1966а, б, 1989 и по пеляди – Андрияшева и др., 1978,1983а; Андрияшева, 1985,1989б), но и за рубежом. - Представление о величине наследуемости различных количественных признаков у рыб дает анализ первых обзорных работ — наиболее полной сводки В.С. Кирпичникова (Kirpichnikov, 1981), содержащей данные по 15 видам рыб из 48 литературных источников, а также обзоров Т. Гьедрема (Gjedrem, 1983, 1986), объединяющих сведения 27 экспериментальных работ по 5 видам рыб, и статьи Б. Кингхорна (Kinghorn, 1983), где помещены материалы по 4 видам из 12 литературных источников. Воспользовавшись этими данными, мы выделили из них для обобщения только те расчеты наследуемости, которые были получены при дисперсионном анализе сибсов или полусибсов «по самцам», как . наиболее точные. Принимая во внимание сходную величину наследуемости одинаковых признаков у разных видов рыб, для усреднения этих показателей использовали наиболее многочисленные данные, полученные при исследовании трех самых массовых объектов разведения (табл. 1.2).
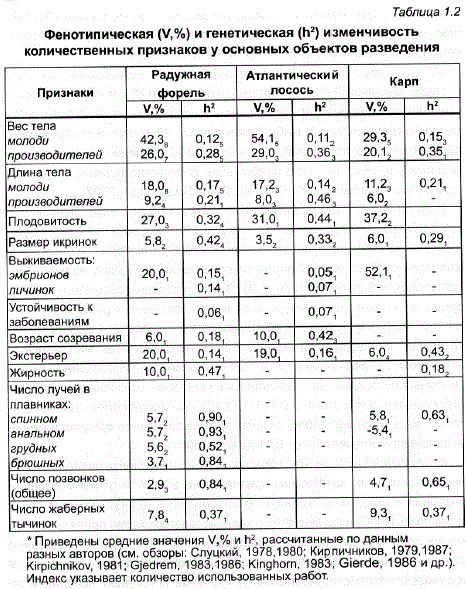
Как видно из табл. 1.2, в целом, несмотря на определенные различия оценок разных авторов, по величине ?? исследованные признаки можно подразделить на две основные группы. Так, у всех трех видов рыб наследуемость была минимальной (0-0,2) при исследовании жизнеспособности икры и личинок, устойчивости к заболеваниям, веса и длины тела в раннем возрасте. У половозрелых рыб уровень генетической изменчи вости по размерно-весовым признакам оказался немного выше (0,3-0,4), чем у молоди. Наиболее значительные показатели наследуемости получены при изучении меристических признаков (0,5-0,9).
Интенсивное развитие какаквакультуры, таки селекционно-генетических исследований в этой области привело в последние 20 лет к многократному увеличению информации, касающейся популяционно-генетических параметров различных видов рыб. Если в первые 10-15 лет были проведены десятки таких исследований, то к настоящему времени они исчисляются многими сотнями, при этом наибольшее количество посвящено различным видам лососевых рыб (Gjedrem, 2000; Garcia et al., 2007).
В наиболее полном к настоящему времени обзоре опубликованы результаты свыше 2000 расчетов наследуемости (в узком смысле) по данным 164 источников (Carlson, Seamons, 2008), в которых исследованы атлантический лосось и 7 видов тихоокеанских лососевых из родов OnCorhynchus и Salvelinus (табл. 1.3). В этой таблице основные признаки, представляющие экономическую ценность, отнесены к различным категориям, каждая из которых характеризуется определенной величиной показателей наследуемости входящих в нее признаков. Так, минимальные значения h? (в среднем от 0,06 до 0,37) выявлены при исследовании практически всех признаков, относящихся к категории «жизненный цикл — life history traits», а максимальные (0,25-0,40) — по морфологическим показателям; категория «физиологических признаков» имела средние и менее варьирующие значения – от 0,11 до 0,19 (рис. 1.1).
Как отмечают авторы этого обзора, полученные оценки наследуемости по определенным категориям признаков не зависели от рода: средние значения h? оказались сходными y Oncorhynchus (0,24), Salvelinus (0,27) и Salmo (0,16). При оценке этих результатов необходимо принять во внимание то, что подавляющее большинство получено при исследовании стад и популяций, обитающих в контролируемых условиях фермерских и заводских хозяйств. Аналогичное изучение природных популяций по объективным причинам проводится очень редко и в данном обзоре (Carlson, Seamons, 2008) составило около 2% от общего числа подсчетов (58 из 2389). Вопреки ожиданиям, величина наследуемости в природных популяциях оказалась несколько ниже, чем в фермерских стадах.
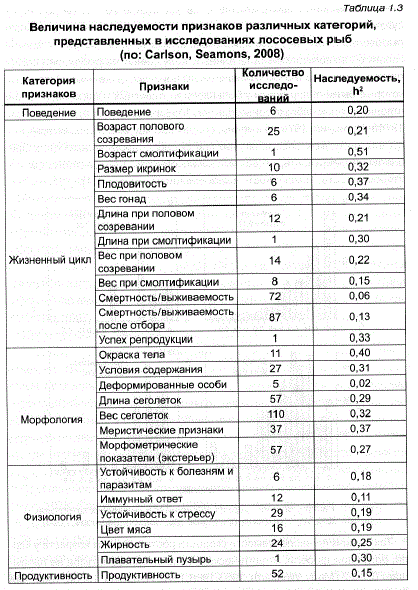
Сравнение значений наследуемости, представленных в табл. 1.2 и 1.3, может свидетельствовать о том, что многократное увеличение количества выполненных расчетов h? за последние 20 лет, не привело к существенным изменениям величины этих показателей по основным рыбоводно-биологическим признакам. Кроме того, несмотря на сравнительную немногочисленность данных, полученных на рыбах, они позволяют считать, что в этом отношении им присущи те же закономерности, которые были обнаружены у большинства животных – как сельскохозяйственных, так и лабораторных (Lerner, 1958; Falconer, 1960; Кирпичников, 1974б; Рокицкий, 1974), а именно: слабая наследуемость характерна для группы признаков, определяющих приспособленность, — таких как эмбриональная смертность, общая жизнеспособность, плодовитость, ранняя скорость роста, устойчивость к заболеваниям (см рис. 1.1).

Так, величина наследуемости скорости роста в раннем возрасте у карпа, радужной форели, атлантического лосося, тиляпии, гуппи не превышает 0,1-0,3 (Moav, Wohlfarth, 1963,1968,1976; Ненашев, 1966а, 1969; Чай Май Тхиен, 1971a; Aulstad et al., 1972; Kincaid, 1972, 1977; Ryman, 1972а, 1973; Chevаssus, 1976; Moller et al., 1979; Refstie, 1980; Tave, Smitherman, 1980; McKay et al., 1986 и др.). Доля генетической изменчивости в варьировании величины плодовитости у радужной форели составляет 0,2-0,4 (Gall, Gross, 1976; Gierde, 1986). Наследуемость смертности икры, личинок и мальков у лосося, кумки и форели не превышает 0,1 (Kanis et al., 1976; McKay et al., 1986), у молоди атлантического лосося равняется 0,1-0,2 (Ryman, 19726). Лишь немного отличаются от нуля значения наследуемости таких признаков, как устойчивость к заболеваниям и неблагоприятным воздействиям лосося, чавычи и озерной форели (Gjedrem, Aulstad, 1974; Сrаmеr, McIntyre, 1975; Gjedrem, 1976а).
Наследуемость другой группы признаков — морфологических и некоторых конституционных – значительно выше – от 0,5 до 0,9. Генетическая изменчивость признаков, имеющих диагностическое значение для многих видов рыб, составляет: для числа позвонков – 0,7-0,9, количества жаберных тычинок – 0,5-0,8, числа лучей в плавниках – 0,4-0,6 (Svardson, 1950,1952; Ненашев, 1966б; Hagen, 1973; Ali, Lindsey, 1974; Кирпичников, 1979; Tave, 1984а, б; Learу et al., 1985а). Показатели наследуемости некоторых экстерьерных и различных счетных признаков у карпа были значительно выше селекционных – 0,4-0,8 (Ненашев, 1966б, 1968; Wohlfarth, 1967). - Наследуемость некоторых экстерьерных, а также физиологичеСких признаков имеет неодинаковые значения у разных рыбоводных объектов. Это может быть связано как со специфическими особенностями биологии вида, так и с различной экономической значимостью рыбоводно-биологических признаков, определяющей направления селекции. Например, у форели и атлантического лосося экстерьерные показатели наследуются слабо — 0,14-0,16 (Gjerde, Gjedrem, 1984), а у карпа, где экстерьер определяет различия между породами, эти значения значительно выше – 0,42-0,49 (Ненашев, 1966б, 1969; Moav, Wohlfarth, 1976). Сложность жизненного цикла у проходных лососевых рыб (Schaffеr, Elson, 1975; Saunders, Schom, 1985) может быть причиной высокой наследуемости возраста полового созревания атлантического лосося – от 0,39 до 1,0 (Naevdal et al., 1976; Gjerde, 1984; Gjerde, Gjedrem, 1984), в отличие от аналогичных показателей у радужной форели — 0,21 (Gjerde, Gjedrem, 1984). В то же время наследуемость жирности у радужной форели довольно высока – 0,47 (Kinghorn, 1981), в два раза выше, чем у карпа, — 0,14-0,22 (Ненашев, 1969; Смишек, 1978; Smisek, 1979).
Этот краткий обзор исследований, посвященных определению уровня генетической изменчивости рыб по количественным признакам, дает представление о перспективности многих экономически важных показателей для селекционной работы. Согласно теоретическим расчетам, на основании полученных значений h? для молоди форели можно ожидать увеличения скорости роста за одно поколение примерно на 7-8% (Gjedrem, 1975; Skjervоld, 1976), а при содержании атлантического поСося в садках – на 2,7-3,6% (Gjedrет, 1983). Сведения об уровне генетической изменчивости основных рыбоводно-биологических признаков определяют стратегию многих селекционных программ (Moav, Wohlfarth, 1960, 1968; Gjedrem, 1976б, 1983; Skjervоld, 1976; Кирпичников, 1979; Wilkins, 1981 и др.).
С другой стороны, результаты этих исследований с несомненностью Свидетельствуют о биологическом смысле показателей наследуемости, В частности, их различий для разных признаков и сходства одних и тех же признаков у разных видов рыб. На это же могут указывать сформированные в процессе эволюции меж- и внутрипопуляционные различия по многим хорошо наследуемым признакам у самых разных видов рыб. Так, Возраст полового созревания атлантического лосося наследственно закреплен не только между крупными стадами (Gunnes, 1978; Naevdal et al., 1978а, б; Gjerde, Refstie, 1984), но и между семьями одного стада (Naevdal, 1983). Генетические различия по сроку нереста можно обнаружить у сигов (Todd, 1981; Vuorinen et al., 1981), тихоокеанских лососей (Алтухов, 1974,1983), радужной и озерной форели, лосося Кларка, разных видов гольцов (Kincaid, 1981). Очевидно, аналогичный механизм определяет различия между популяциями по размеру икринок (Волохонская, Викторовский, 1971; Aulstad, Gjedrem, 1973; Слуцкий, 1980), а также по величине меристических признаков – количеству позвонков, жаберных тычинок, числу лучей в плавниках (см.: Svardsen, 1952; Кирпичников, 1979; Решетников, 1980).
В то же время при изучении природных популяций редко удается выявить генетические различия между ними по признакам приспособленности. В подавляющем большинстве случаев дифференциация стад или популяций по скорости роста, плодовитости, устойчивости к заболеваниям является следствием интенсивной и длительной селекционной работы. Таким образом, закономерность, отмеченная Д. Фолкнером (1985, с. 147), — степень сходства потомков с родителями, измеряемая наследуемостью, — «меняется вместе с признаками, по некоторым она больше, по другим меньше», в четкой форме проявляется и при исследований генетической изменчивости количественных признаков в природных и одомашненных популяциях рыб.
Не менее существенными при исследовании рыб оказались различия в уровне изменчивости различных экономически важных показателей, при этом по степени вариабельности их также удалось разделить на группы. Согласно классификации, предложенной Е.С. Слуцким (1978), уровень изменчивости количественных признаков у рыб в зависимости от величины коэффициента изменчивости может быть оценен как низкий (до 5%), средний (10-20%), сильный (20-40%) и высокий (40-50% и более). Как выявило это исследование, у рыб слабо варьирующими и относительно менее зависимыми от условий среды оказались все меристические признаки, размер икринок и некоторые экстерьерные индексы. Распределение таких признаков подчиняется нормальному закону, а коэффициенты изменчивости обычно не превышают 1-7% (Правдин, 1929; Чан Май Тхиен, 1971б; Канеп, 1973; Слуцкий, Тищенко, 1973а; Слуцкий, 1978,1980; Тищенко, 1978б; Learу et al., 1985а и др.).
К группе признаков, отличающихся высокой вариабельностью, принадлежат те, которые имеют адаптивное значение, — вес тела в раннем возрасте, плодовитость, жизнеспособность. Они чрезвычайно подвержены средовым влияниям, часто характеризуются асимметричным распределением и обладают высокими коэффициентами вариации – 20-50% и более (Кирпичников, 1974б, 1979; Gjedrem, 1975, 1979, 1983; Слуцкий, 1978). Таким образом, даже при этом кратком обзоре исследований вариабельности количественных признаков у рыб удается обнаружить те же закономерности, которые наблюдаются и у других животных (Lerner, 1958; Falconer, 1960; Рокицкий, 1974 и др.).
К аналогичному выводу приводит и сопоставление уровня фенотипической и генетической изменчивости различных мерных признаков у разных видов рыб (см. табл. 1.2). Оно свидетельствует о том, что у рыб, как и у других животных, между показателями наследуемости и коэффициентами вариации существует обратная зависимость; при высоком уровне вариабельности признака (20-50%) его селекционная ценность обычно невелика (< 0,3), тогда как признаки, отличающиеся пониженной фенотипической изменчивостью (до 10%), имеют в большинстве случаев высокую наследственную обусловленность (0,5-0,9). Весьма вероятно, что дальнейшие исследования в этом направлении позволят уточнить подобные представления. В настоящее время выявленная зависимость, По-видимому, может быть использована в практической селекции рыб для самой первой, предварительной оценки природы различных хозяйственно полезных признаков.