Влияние освещенности на рост и развитие личинок нельмы
Как известно, солнечный свет как абиотический фактор играет важную роль в жизни большинства видов рыб практически на всех этапах развития. Особенно значимым является влияние освещенности на ранние стадии постэмбриогенеза, когда успех выживания молоди во многом определяет динамику численности популяций.
Публикаций о влиянии света на рыб довольно много, но в большинстве из них рассматриваются вопросы поведения, двигательной активности, размножения и питания (Лещева, 1967; Волкова, 1971, 1973; Гирса, 1981; Савченко и др., 1981).
Вместе с тем, особый интерес представляют данные о воздействии света на процессы роста и выживаемость молоди. В литературе этот вопрос освещен достаточно полно на разных видах рыб (Barlow et al., 1995; Gulbrandsen et al., 1996; Abdel-Fattah, Mamdouh, 2004; Гутиева 2005; Ручин, 2006), однако представители семейства сиговых остаются малоизученными. Работы о влиянии светового фактора на личиночное развитие сиговых рыб в основном направлены на поиск оптимальных режимов освещенности при подращивании молоди в искусственных условиях (Раденко, Терентьев, 1988; Решетников и др., 1989).
Изучение влияния различного уровня освещенности на развитие, выживаемость и поведение личинок кубенской нельмы проводились нами в двух прямоточных лотках с идентичными условиями. Контрольную группу личинок разместили в лотке, находившемся в помещении рыбоводного цеха, опытную - в лотке, установленном на открытой площадке вне цеха. Опытный лоток был накрыт тентом, покрывающим около 75% водной поверхности между местом подачи воды и сливным устройством. На момент начала эксперимента личинки нельмы в возрасте 28 сут. после вылупления имели среднюю длину 19,4 мм и массу 29,5 мг. Желточный мешок был израсходован, личинки находились на III этапе развития. Подробная характеристика личиночного развития нельмы приведена ранее (см. главу 4.2).
Начальная плотность посадки в обоих лотках равнялась 20 тыс. экз./лоток (17 тыс./м3). Нормы кормления рассчитывали в соответствии с методическими указаниями по выращиванию сиговых рыб (Костюничев и др., 2005); использовали искусственные корма ?Troco? (?Coppens?, Голландия).
Продолжительность эксперимента составила 29 сут. (1-29 июня), температура воды в этот период находилась в пределах 13,9-16,2°C. Освещенность лотков приведена в таблице 14.
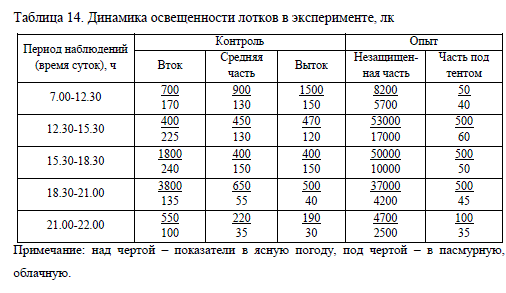
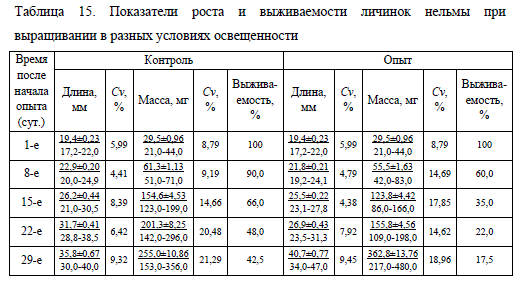
Результаты исследований показали, что высокая интенсивность освещения на ранних этапах постэмбрионального развития крайне отрицательно влияет на выживаемость и рост личинок нельмы. В течение первых 7 сут. после рассаживания молоди в опыте наблюдалась повышенная смертность, которая составила около 40% исходного числа личинок против 10% в контроле (табл. 15).
Высокая смертность подопытной нельмы может быть связана с ее поведением – после пересадки в условия высокой инсоляции молодь не укрывалась в затененных тентом участках рыбоводной емкости, а равномерно распределялась по всей ее площади.
В последующем динамика смертности нельмы в двух группах существенно не различалась, а элиминация нельмы в эксперименте была связана с физиологическими изменениями организма молоди, такими как появление плавательного пузыря, начало функционирования желудка и др.
Через неделю после начала эксперимента у личинок в опыте было отмечено отставание в развитии: большинство из них (68%) находились на IV этапе личиночного развития (наполнение плавательного пузыря воздухом, окостенение лучей в плавниках, обособление желудка); всего 28% молоди перешли на V этап (появление в перитонеуме серебристого пигмента гуанина, хвостовой плавник начинает принимать гомоцеркальную форму). В контроле в это время большинство особей (64%) находились на V этапе развития. В обоих вариантах эксперимента наблюдались особи (по 4% в каждом), оставшиеся на III этапе, их плавательный пузырь не был наполнен воздухом. При вскрытии и исследовании этих личинок с помощью бинокулярного микроскопа было обнаружено патологическое перекрытие канала, соединяющего плавательный пузырь с кишечным трактом.
Дальнейшее развитие личинок нельмы, до их перехода на мальковый этап, связано с формированием петлеобразного изогнутого желудка. Эта стадия у большинства личинок наблюдалась на 15-е сут. эксперимента (в контроле – 64%, в опыте – 52%), а ее продолжительность составляла 10 сут. (с 13 по 22 июня). За неделю до окончания опыта (22-е сут.) в контроле доминировали особи, находившиеся на стадии перехода от личиночного к мальковому периоду. В опыте такой молоди было значительно меньше. Однако к окончанию эксперимента практически вся молодь в опыте и контроле имела признаки малькового развития – появление зачатков чешуи.
Данные таблицы 15 свидетельствуют о том, что в течение почти всего эксперимента подопытная молодь росла медленнее, чем в контроле, масса личинок в опыте была меньше на 23% по сравнению с массой контрольных рыб. На них оказывало негативное влияние избыточное количество света, интенсивность которого на водной поверхности рыбоводной емкости, не защищенной тентом, иногда достигала 53000 лк, в то время как на поверхности лотка в помещении этот показатель не превышал 4000 лк (табл. 14). Увеличение конечной массы подопытных рыб связано с резким снижением плотности посадки из-за высокой смертности.
После перехода молоди нельмы на мальковый период развития смертность в обеих группах практически прекратилась. Выживаемость личинок в контроле составила 42,5% от числа личинок в начале эксперимента (8500 экз.), а в опыте – 17,5% (3500 экз.). По достижении молодью малькового этапа эксперимент был прекращен, а нельма высажена в садок, расположенный в озере. Мальки держались на глубине, поднимаясь на поверхность исключительно во время кормления.
Высокую смертность личинок, которая наблюдалась с первой недели эксперимента в опытном варианте, можно объяснить рядом причин. С одной стороны, это – стресс, вызванный сменой условий содержания, с другой – слабая пигментация поверхности тела и отсутствие гуанинового пигмента в перитонеуме молоди (он образуется позже и наряду с пигментацией обеспечивает резистентность рыб к солнечной радиации). Отсутствие защиты от чрезмерной инсоляции негативно сказывалось на росте и выживаемости молоди нельмы в новых условиях после пересадки, так как личинки игнорировали затемненные участки водной поверхности и равномерно рассредоточивались по всему объему рыбоводной емкости. Следует подчеркнуть, что в течение первых 10 сут. опыта преобладала ясная солнечная погода.
На протяжении всего эксперимента личинки в обеих группах различались по размерным показателям. Относительно высокий прирост массы и увеличение линейных размеров молоди в опыте был отмечен на последнем этапе личиночного развития. К этому времени число рыб в лотке существенно уменьшилось в связи с элиминацией отстающих особей, что определило более благоприятные условия для дальнейшего развития оставшейся молоди. Тем не менее, избыточная освещенность негативно влияет на сроки наступления отдельных этапов личиночного развития, скорость их прохождения, темп роста и выживаемость нельмы. При чрезмерном воздействии солнечной инсоляции наблюдалась задержка роста и отставание в развитии личинок нельмы в опытной группе. В ней смертность личинок за период эксперимента составила 82,5%, что в 1,4 раза выше, чем в контроле (57,5%).
Схожие выводы были сделаны А.Б. Ручиным (2009) в эксперименте на сеголетках четырех видов рыб (серебряного карася Carassius auratus gibelio, золотого карася C. carassius, ротана Perccottus glenii и сибирского осетра Acipenser baerii) при выдерживании их в диапазоне освещенности 0–9000 лк.
Автор показал, что скорость роста молоди снижалась по мере увеличения интенсивности света. По данным Е.Л. Мельниковой (1986), у личинок атлантического лосося Salmo salar при повышении освещенности до 500 лк наблюдается снижение показателей выживаемости, среднесуточных приростов длины и массы тела, а также замедление резорбции желтка и задержка перехода на активное питание.
Интересные результаты были получены при исследовании влияния разных световых режимов на поведение сиговых. В частности, в экспериментах В.Н. Раденко и П.В. Терентьева (1988) не было обнаружено отклонений в поведении личинок пеляди, которых содержали в довольно широком диапазоне освещенности – 320-20000 лк. Аналогичные результаты были получены и в наших исследованиях: изменение интенсивности освещения не сопровождалось изменением поведения молоди нельмы; личинки не избегали участков бассейна, подверженных высокой инсоляции.
Данные о влиянии на личинок рыб высокой степени освещенности и воздействия солнечного света в доступной литературе немногочисленны. Например, С.Г. Крыжановский с соавторами (1953) на личинках судака Sander lucioperca показали, что сильная освещенность может привести к высокой смертности рыб, особенно чувствительны личинки на ранних этапах развития.
Воздействие прямого солнечного света на личинок этого вида приводит к их гибели в течение 3-4 ч (Woynarovich, 1960). Е. Войнарович предположил, что смерть личинок могла наступать по причине ослепления и поражение участков нервной системы, отвечающих за движение и равновесие. Зарубежными исследователями Л. Кораццей и С. Никумом (Corazza, Nickum, 1981 – цит. по: Королеву, 1999) было установлено, что высокая освещенность блокирует пищевой рефлекс у американского судака Stizostedion vitreum.
Повреждающее действие прямого светового излучения на живые организмы было описано в работе Д.И. Рощупкина и А.Я. Потапенко (1977). Оно заключается в нарушении структуры и функций клеток и клеточных образований, инактивации некоторых белков и нуклеиновых кислот, фотоповреждении клеточных структур. Позже, эти данные были подтверждены В.Н. Жукинским (1986) на различных видах рыб.
Таким образом, можно предположить, что даже кратковременное прямое воздействие избыточного количества солнечной инсоляции способно нанести вред молоди рыб, особенно на ранних этапах личиночного развития, когда защитные механизмы от ультрафиолетового излучения развиты слабо.
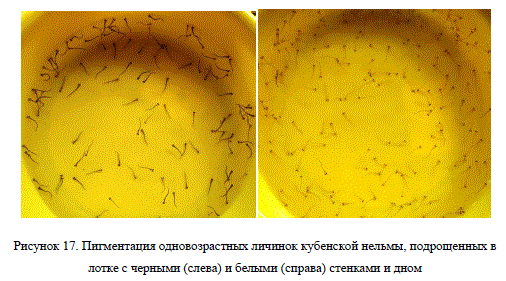
Интенсивной пигментации тела личинок нельмы можно добиться при их выращивании в лотках с темными стенками и дном. Наблюдения показали, что использование лотков черного цвета позволяет получить намного более пигментированную молодь, чем при выращивании нельмы в белых лотках (рис. 17). К сожалению, оценить резистентность такой пигментации к солнечной освещенности нам не удалось. Но, следует полагать, что сильно пигментированные личинки менее восприимчивы к воздействию чрезмерной инсоляции, т. к. меланофоры у рыб позволяют регулировать светопропускание через кожу (Микулин, 2000).
На основании проведенных исследований по влиянию солнечной инсоляции на личинок нельмы при выращивании в индустриальных условиях можно рекомендовать использование затенения всей рыбоводной емкости до момента обретения молодью резистентности в виде образования в теле гуанинового пигмента. Возраст личинок нельмы к этому времени составляет около 40 сут.