9.5. Особенности влияния солнца на развитие сигов
Исследованиями многих отечественных и зарубежных ихтиологов установлено, что
световая энергия суммарно действует на зародышевое развитие рыб подобно температу-
ре. Это выражается в том, что повышение уровня освещенности в период эмбриогенеза
вызывает сокращение числа формирующихся туловищных сегментов, позвонков, пробо-
дённых чешуй в боковой линии и количества лучей в непарных плавниках (Smith, 1916;
Scheffelt, 1926; Lindsey, 1958; Рощупкин, Потапенко, 1977). Таким образом, подтверждает-
ся, что «Второе правило Джордана» (1892) в отношении температурного фактора — чем
выше температура развития, тем меньше значения меристических признаков, — приме-
нимо и к воздействию света. Доказано, что солнечная радиация в видимом диапазоне не
менее чем температура, определяет жизнеспособность эмбрионов рыб, темпы развития
и процессы дифференцировки тканей (Viber, 1954; Любицкая, 1961; Любицкая, Дорофее-
ва, 1961; Коровина, Любицкая, Дорофеева, 1965).
С. Шеффельт (Scheffelt, 1926), исследуя воздействие света на эмбриональное разви-
тие сиговых рыб, отметил, что увеличение уровня освещенности вызывает снижение
числа позвонков и других меристических признаков. Это подтвердили работы К. Джона
и А. Хаслера (John, Hasler, 1956) о влиянии светового фактора на эмбриогенез и выжи-
ваемость икры сиговых рыб озера Мендота. Проводя опыты по воздействию солнечной
радиации на эмбриогенез озёрного сига (Coregonus wartmanni Bloch), немецкий учёный
Е. Браум (Braum, 1964), констатировал, что солнечная радиация в видимом диапазоне
положительно влияет на выживаемость икры сиговых рыб и ускоряет темп развития
и вылупления личинок из оболочки.
Исследования оптических характеристик икры омуля при помощи спектрофотометра
показали (Черняев и др., 1988; Валюшок и др., 1993), что поглощающая способность икры
в различных участках спектра по мере её развития возрастает. Наибольшая оптическая
плотность икры находится в синей и фиолетовой областях видимого диапазона спектра
(300 нм), наименьшая — в желтой (450 нм) и красной (500 нм). Икра омуля (и других
сиговых рыб) окрашена содержащимися в ней каротиноидами и цитохромом в светло-о-
ранжевый цвет (600–580 нм). Зеленый (520 нм), синий (470 нм) и фиолетовый (460 нм)
цвета спектра проникающей под лёд в воду световой радиации являются дополнительны-
ми к оранжевому и жёлтому, что способствует более полному поглощению зародышем
сиговой рыбы лучистой энергии солнечного света.
В осенние месяцы развития, когда в теле зародышей осенне-нерестующего омуля еще
не сформированы специализированные органы фоторецепции (и, главным образом, от-
сутствуют миотомы), степень освещенности практически не оказывает влияния на темп
эмбриогенеза, как в природных, так и в искусственных условиях (Черняев, 1993, 2007).
К декабрю на этапе органогенеза, с начала сегментации мезодермы в теле эмбриона
формируются нервная трубка, кишечный тяж, хорда и другие осевые органы. В форми-
рующихся мышечных сегментах происходит интенсивная пролиферация (размножение)
клеток, сопровождающаяся выработкой нейротрасмиттеров, которые совместно с ней-
ропептидами (энкефалинами) в переданной самкой яйцеклетке воздействуют на темп
176
и последовательность морфогенетических процессов (Бузников, 1967, 1987, Макеева,
1992, Черняев, 2007). Позднее появляются другие более сложно организованные фото-
чувствительные образования: в теменном отделе головного мозга — эпифиз, а по бокам
головы — глазные бокалы. На желточном мешке, вдоль кишечной трубки и тела зароды-
ша появляются меланофоры, а в сосудах эмбрионального кровообращения формируются
окрашенные гемоглобином форменные элементы крови.
С февраля, по мере возрастания уровня инсоляции, всё в большей степени проявляется
воздействие солнечной радиации на эмбриогенез омуля. В результате этого вылупление
эмбрионов на естественных нерестилищах происходит в апреле при низкой температуре
воды (1,7° C) сразу после ледохода, а инкубируемая на рыбоводном заводе икра омуля
в условиях низкой освещенности продолжает развиваться до конца мая. Вылупление
эмбрионов происходит за счет весеннего прогрева речной воды (7,5° C), поступающей
в инкубационный цех (Черняев, Довгий, 1969, Черняев, 1982, 1984, 2010).
Ниже приведены результаты измерений солнечной радиации, проходящей под лед
и достигающей поверхности нерестилищ, проведенные с помощью подводного пирано-
метра системы Янишевского, оснащённого черно-белой термобатареей площадью термо-
пар 7?7 см и «зайчиковым» высокочувствительным гальванометром (Черняев, Довгий,
1969) (табл. 27).
Эти исследования проводились в исключительно неблагоприятных для прохождения
света условиях — лёд над нерестилищами омуля был мутным, пористым, многослой-
ным. Между слоями льда в результате ледяного затора реки образовалась «шуга», затем
в середине зимы появились наледи. Общая глубина от поверхности льда до дна была
около 180 см, а поверхность льда покрыта снегом толщиной от 40 до 60 см. При условиях
прозрачного кристаллического льда, через который солнечный свет проходит практиче-
ски беспрепятственно, показатели освещенности у дна были бы значительно выше. Тем
не менее, даже в столь неблагоприятных для прохождения в воду солнечной радиации
до дна нерестилищ условиях, зафиксированные значения поглощённой световой энергии
в природных условиях (5851,7 Дж/10–4 на м2 или 1396,6 кал/см2) многократно превосхо-
дили суммарные значения солнечной энергии, получаемой икрой в условиях искусствен-
ного разведения (Черняев, Довгий, 1969).
Таблица 27.
Количество световой энергии, прошедшей под лед реки Большой в 1964–65 г.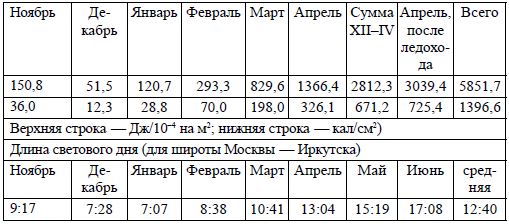
Замеры световой радиации в цехе Большереченского рыбоводного завода на наружной
поверхности и внутри инкубационных аппаратов Вейса, содержащих по 250000 икринок
омуля, показали, что за время инкубации (210–220 cуток) на поверхность аппарата при
естественном освещении солнечным светом поступило 251,4 Дж/10–4/м2 или 60 кал/см2,
что в 23,3 раза меньше, чем получила икра на нерестилище (Черняев, 1982, 1990). При
этом за счет процесса перемешивания икры в аппарате Вейса количество световой энер-
гии, доступное для каждого эмбриона, составило лишь незначительную часть (в среднем
6,5%) от поступающей к инкубационным аппаратам! Водоснабжение стоек с аппаратами
Вейса на заводе осуществляется непосредственно с нерестилищ реки Большой, что обе-
спечивает естественный температурный режим инкубации икры. Икра в цехах рыбово-
дного предприятия, недополучая солнечную энергию, развивается на полтора-два месяца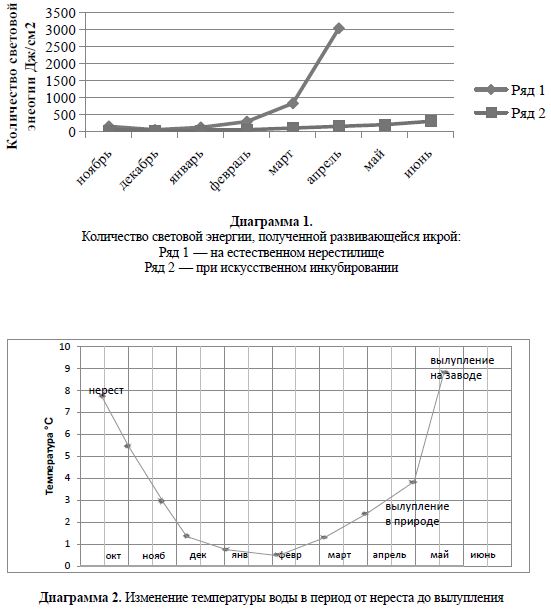
дольше, и вылупление эмбрионов происходит только при значительном повышении тем-
пературы воды, закачиваемой из реки (Черняев, 2010). Согласно расчетам К. И. Мишарина
(1953), за счет удлинения сроков инкубации, при весеннем прогреве речной воды, икра на
рыбоводных заводах дополнительно получает 20% тепла, которые очевидно восполняют
недостаток солнечной энергии, необходимой для завершения эмбриогенеза и вылупления
эмбриона из оболочки.
Диаграммы 1 и 2 отображают динамику температуры и светового излучения в период
эмбрионального развития байкальского омуля как в естественных условиях нерестилищ,
так и в цехах Большереченского рыбоводного завода.
Диаграммы позволяют вычленить приоритет светового фактора над температурным
(по разнице поглощённой световой энергии) при одинаковой температуре как в есте-
ственных условиях развития икры на нерестилищах, так и при искусственной инкубации.
То есть для осенненерестующих рыб на поздних этапах эмбриогенеза ведущим фактором
воздействия на темп развития и сроки вылупления является световой, а не температур-
ный.
Оптическая плотность икры по мере развития эмбриона возрастает. Так, на этапах
дробления и гаструляции (рис. 70) икра омуля поглощает в среднем 30,1%, на этапе ор-
ганогенеза — 53,8%, а на последующих этапах развития вплоть до вылупления порядка
76,4% поступающей радиации. Измерения проводились при помощи спектрофотометров
СФ?4а и СФ?10 (Черняев, Довгий, 1969).

Рис. 70. Изменение оптической плотности икры D%
1 — температура среды; 2 — оптическая плотность икры
179
В работах Т. Линдстрема (Lindstrёm, 1970) по исследованию условий размножения
и развития сиговых рыб Швеции показано, что температура и освещённость в реках
и озёрах значительно отличаются, а энергия солнца ускоряет и регулирует темп разви-
тия икры сиговых рыб. Немецкими рыбоводами (Кох и др., 1980) установлено, что икра
лососеобразных рыб — семги (Salmo salar), кумжи (Salmo trutta), ручьевой форели
(Salmo trutta trutta), дунайского тайменя (Hucho taimen), гольца (Salvelinus alpinus), палии
(Salvelinus lepechini) и хариуса (Thymallus thymallus) в заводских условиях под влиянием
солнечной радиации ускоряет свое развитие, но увеличивается и количество аномально
развитых зародышей.